
This report contains the collective views of an international group of experts and does not necessarily represent the decisions or the stated policy of the United Nations Environment Programme, the International Labour Organization, or the World Health Organization.
Concise International Chemical Assessment Document 57
First draft prepared by Drs J. Kielhorn, C. Pohlenz-Michel, S. Schmidt, and I. Mangelsdorf, Fraunhofer Institute for Toxicology and Experimental Medicine, Hanover, Germany
Published under the joint sponsorship of the United Nations Environment Programme, the International Labour Organization, and the World Health Organization, and produced within the framework of the Inter-Organization Programme for the Sound Management of Chemicals.
World Health Organization
Geneva, 2004
The International Programme on Chemical Safety (IPCS), established in 1980, is a joint venture of the United Nations Environment Programme (UNEP), the International Labour Organization (ILO), and the World Health Organization (WHO). The overall objectives of the IPCS are to establish the scientific basis for assessment of the risk to human health and the environment from exposure to chemicals, through international peer review processes, as a prerequisite for the promotion of chemical safety, and to provide technical assistance in strengthening national capacities for the sound management of chemicals.
The Inter-Organization Programme for the Sound Management of Chemicals (IOMC) was established in 1995 by UNEP, ILO, the Food and Agriculture Organization of the United Nations, WHO, the United Nations Industrial Development Organization, the United Nations Institute for Training and Research, and the Organisation for Economic Co-operation and Development (Participating Organizations), following recommendations made by the 1992 UN Conference on Environment and Development to strengthen cooperation and increase coordination in the field of chemical safety. The purpose of the IOMC is to promote coordination of the policies and activities pursued by the Participating Organizations, jointly or separately, to achieve the sound management of chemicals in relation to human health and the environment.
WHO Library Cataloguing-in-Publication Data
Glyoxal.
(Concise international chemical assessment document ; 57)
1.Glyoxal — toxicity 2.Risk assessment 3.Environmental exposure 4. Food contamination
I.International Programme on Chemical Safety II.Series
ISBN 92 4 153057 X (LC/NLM Classification: QD 305.A6)
ISSN 1020-6167
©World Health Organization 2004
All rights reserved. Publications of the World Health Organization can be obtained from Marketing and Dissemination, World Health Organization, 20 Avenue Appia, 1211 Geneva 27, Switzerland (tel: +41 22 791 2476; fax: +41 22 791 4857; email: bookorders@who.int). Requests for permission to reproduce or translate WHO publications — whether for sale or for noncommercial distribution — should be addressed to Publications, at the above address (fax: +41 22 791 4806; email: permissions@who.int).
The designations employed and the presentation of the material in this publication do not imply the expression of any opinion whatsoever on the part of the World Health Organization concerning the legal status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. Dotted lines on maps represent approximate border lines for which there may not yet be full agreement.
The mention of specific companies or of certain manufacturers’ products does not imply that they are endorsed or recommended by the World Health Organization in preference to others of a similar nature that are not mentioned. Errors and omissions excepted, the names of proprietary products are distinguished by initial capital letters.
The World Health Organization does not warrant that the information contained in this publication is complete and correct and shall not be liable for any damages incurred as a result of its use.
Risk assessment activities of the International Programme on Chemical Safety, including the production of Concise International Chemical Assessment Documents, are supported financially by the Department of Health and Department for Environment, Food & Rural Affairs, UK, Environmental Protection Agency, Food and Drug Administration, and National Institute of Environmental Health Sciences, USA, European Commission, German Federal Ministry of Environment, Nature Conservation and Nuclear Safety, Health Canada, Japanese Ministry of Health, Labour and Welfare, and the Swiss Agency for Environment, Forests and Landscape.
Technically and linguistically edited by Marla Sheffer, Ottawa, Canada, and printed by Wissenchaftliche Verlagsgesellschaft mbH, Stuttgart, Germany
Concise International Chemical Assessment Documents (CICADs) are the latest in a family of publications from the International Programme on Chemical Safety (IPCS) — a cooperative programme of the World Health Organization (WHO), the International Labour Organization (ILO), and the United Nations Environment Programme (UNEP). CICADs join the Environmental Health Criteria documents (EHCs) as authoritative documents on the risk assessment of chemicals.
International Chemical Safety Cards on the relevant chemical(s) are attached at the end of the CICAD, to provide the reader with concise information on the protection of human health and on emergency action. They are produced in a separate peer-reviewed procedure at IPCS. They may be complemented by information from IPCS Poison Information Monographs (PIM), similarly produced separately from the CICAD process.
CICADs are concise documents that provide summaries of the relevant scientific information concerning the potential effects of chemicals upon human health and/or the environment. They are usually based on selected national or regional evaluation documents or on existing EHCs. Before acceptance for publication as CICADs by IPCS, these documents undergo extensive peer review by internationally selected experts to ensure their completeness, accuracy in the way in which the original data are represented, and the validity of the conclusions drawn.
The primary objective of CICADs is characterization of hazard and dose-response from exposure to a chemical. CICADs are not a summary of all available data on a particular chemical; rather, they include only that information considered critical for characterization of the risk posed by the chemical. The critical studies are, however, presented in sufficient detail to support the conclusions drawn. For additional information, the reader should consult the identified source documents upon which the CICAD has been based.
Risks to human health and the environment will vary considerably depending upon the type and extent of exposure. Responsible authorities are strongly encouraged to characterize risk on the basis of locally measured or predicted exposure scenarios. To assist the reader, examples of exposure estimation and risk characterization are provided in CICADs, whenever possible. These examples cannot be considered as representing all possible exposure situations, but are provided as guidance only. The reader is referred to EHC 170.1
While every effort is made to ensure that CICADs represent the current status of knowledge, new information is being developed constantly. Unless otherwise stated, CICADs are based on a search of the scientific literature to the date shown in the executive summary. In the event that a reader becomes aware of new information that would change the conclusions drawn in a CICAD, the reader is requested to contact IPCS to inform it of the new information.
Procedures
The flow chart on page 2 shows the procedures followed to produce a CICAD. These procedures are designed to take advantage of the expertise that exists around the world — expertise that is required to produce the high-quality evaluations of toxicological, exposure, and other data that are necessary for assessing risks to human health and/or the environment. The IPCS Risk Assessment Steering Group advises the Coordinator, IPCS, on the selection of chemicals for an IPCS risk assessment based on the following criteria:
Thus, it is typical of a priority chemical that
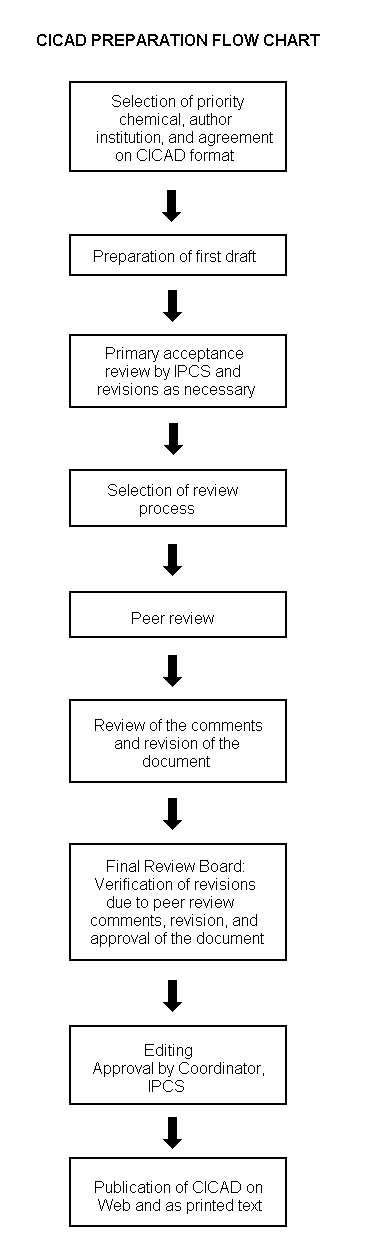
|
Advice from Risk Assessment Steering Group Criteria of priority:
Thus, it is typical of a priority chemical that
Special emphasis is placed on avoiding duplication of effort by WHO and other international organizations. A prerequisite of the production of a CICAD is the availability of a recent high-quality national/regional risk assessment document = source document. The source document and the CICAD may be produced in parallel. If the source document does not contain an environmental section, this may be produced de novo, provided it is not controversial. If no source document is available, IPCS may produce a de novo risk assessment document if the cost is justified. Depending on the complexity and extent of controversy of the issues involved, the steering group may advise on different levels of peer review:
|
The Steering Group will also advise IPCS on the appropriate form of the document (i.e., a standard CICAD or a de novo CICAD) and which institution bears the responsibility of the document production, as well as on the type and extent of the international peer review.
The first draft is usually based on an existing national, regional, or international review. When no appropriate source document is available, a CICAD may be produced de novo. Authors of the first draft are usually, but not necessarily, from the institution that developed the original review. A standard outline has been developed to encourage consistency in form. The first draft undergoes primary review by IPCS to ensure that it meets the specified criteria for CICADs.
The second stage involves international peer review by scientists known for their particular expertise and by scientists selected from an international roster compiled by IPCS through recommendations from IPCS national Contact Points and from IPCS Participating Institutions. Adequate time is allowed for the selected experts to undertake a thorough review. Authors are required to take reviewers’ comments into account and revise their draft, if necessary. The resulting second draft is submitted to a Final Review Board together with the reviewers’ comments. At any stage in the international review process, a consultative group may be necessary to address specific areas of the science. When a CICAD is prepared de novo, a consultative group is normally convened.
The CICAD Final Review Board has several important functions:
Board members serve in their personal capacity, not as representatives of any organization, government, or industry. They are selected because of their expertise in human and environmental toxicology or because of their experience in the regulation of chemicals. Boards are chosen according to the range of expertise required for a meeting and the need for balanced geographic representation.
Board members, authors, reviewers, consultants, and advisers who participate in the preparation of a CICAD are required to declare any real or potential conflict of interest in relation to the subjects under discussion at any stage of the process. Representatives of nongovernmental organizations may be invited to observe the proceedings of the Final Review Board. Observers may participate in Board discussions only at the invitation of the Chairperson, and they may not participate in the final decision-making process.
This CICAD on glyoxal was prepared by the Fraunhofer Institute for Toxicology and Experimental Medicine, Hanover, Germany. It is based on reports compiled by the German Advisory Committee on Existing Chemicals of Environmental Relevance (BUA, 1997). A comprehensive literature search of relevant databases was conducted up to February 2003 to identify any relevant references published subsequent to those incorporated in these reports. Information on the preparation and peer review of the source document is presented in Appendix 1. Information on the peer review of this CICAD is presented in Appendix 2. This CICAD was considered and approved as an international assessment at a meeting of the Final Review Board, held in Varna, Bulgaria, on 8-11 September 2003. Participants at the Final Review Board meeting are presented in Appendix 3. The International Chemical Safety Card for glyoxal (ICSC 1162), produced by the International Programme on Chemical Safety (IPCS, 2002), has also been reproduced in this document.
Anhydrous glyoxal (CAS No.
The predominant target compartments for glyoxal in the environment are the hydrosphere and soil (at about 46% and 54%, respectively) and, to a lesser extent, air (<1%). Reported concentrations of glyoxal in ambient air in the USA, Europe, and Asia are in the range of about 0.1 µg/m3 up to 10 µg/m3. In European rivers and groundwater, concentrations up to 12 µg/litre are reported. Glyoxal is a by-product of ozone disinfection and has been detected at low µg/litre concentrations in drinking-water.
Due to microbial activity as well as non-enzymatic autoxidation of oil or browning reactions of saccharides, glyoxal is frequently detected in fermented food and beverages. It was found in different brands of beer, wine, and other beverages such as tea at concentrations ranging from about 20 µg/litre (black tea) up to 1556 µg/litre (sherry wine). In addition, it was detected in a range of fermented products such as soybean paste and yoghurt (0.63-4.2 mg/kg), bakery products such as bread (0.07-1.6 mg/kg), different plant materials (3-14 mg/kg), and edible oils (up to 6.5 mg/kg).
Glyoxal released into the environment is rapidly converted by abiotic processes, such as transformation by photochemically produced hydroxyl radicals. Due to the low soil sorption coefficient (Koc) reported for this compound, it may leach from soil into groundwater. However, it is readily biodegraded and quickly transformed enzymatically by bacteria and fungi. Its low log octanol/water partition coefficient (Kow) indicates that glyoxal is unlikely to bioaccumulate.
The main routes of occupational exposure to glyoxal during use as a disinfectant are via inhalation of aerosol and dermal absorption. The general population is exposed mainly through ingestion of glyoxal-containing food, but could be exposed through polluted air in urban regions and through traces of glyoxal in drinking-water.
Glyoxal is endogenously produced during normal cellular metabolism by a multitude of enzyme-independent pathways. Glyoxal is also a product of the metabolism and microsomal oxidation of other compounds, such as glycolaldehyde, ethylene glycol, and beta-hydroxy-substituted N-nitrosamines. The concentration of glyoxal in human blood plasma has been reported to be 0.1-1 µmol/litre, with higher levels reported for patients with diabetes or renal failure. In biological materials, less than 10% of the glyoxal present is in unbound forms in aqueous solution (free glyoxal and hydrates), as most of the reactive carbonyl groups are reversibly bound to cysteinyl, lysyl, and arginyl residues of proteins.
Glyoxal, which attacks amino groups of proteins, nucleotides, and lipids, is considered an important intermediate in the formation of advanced glycation end-products (AGEs). AGE modification alters protein function and inactivates enzymes, resulting in disturbance of cellular metabolism, impaired proteolysis, and inhibition of cell proliferation and protein synthesis. The deleterious effects of the highly reactive glyoxal are counteracted by a ubiquitous glutathione (GSH)-dependent glyoxalase system, which converts glyoxal to the less reactive glycolate.
The acute toxicity of glyoxal in experimental animals is low to moderate, depending on the actual concentration of glyoxal in the tested product. In rats, for 40% glyoxal, the LC50 for a single 4-h inhalation of aerosol is 2440 mg/m3, the oral LD50 value ranges from 3000 to 9000 mg/kg body weight (with higher sensitivity in females), and dermal LD50 values are >2000 mg/kg body weight. After inhalation exposure, local irritations of the eyes and respiratory organs as well as hyperaemia and foamy secretion in the lungs predominate. After oral exposure to glyoxal, macroscopic observations include irritations of the gastrointestinal tract and congestion in the gastrointestinal tract, lung, kidney, and adrenal glands. In the prominent target organs, pancreas and kidney, the toxic action of glyoxal leads to severe degenerative changes resembling those induced during diabetes.
Studies into short-term (29-day) inhalation exposure of rats to glyoxal showed a no-observed-effect level (NOEL) of 0.6 mg/m3 (nominal concentration was 0.4 mg/m3) for local effects in the larynx and a NOEL of >8.9 mg/m3 (nominal concentration was 10 mg/m3) for systemic effects (examination of body weight, haematological and biochemical parameters, urine analysis, macroscopic and histological examination). A 28-day study in which glyoxal was administered to rats in drinking-water resulted in a no-observed-adverse-effect level (NOAEL) of 100 mg glyoxal/kg body weight per day. The 90-day feeding of glyoxal to rats resulted in a NOAEL of 125 mg/kg body weight per day (dosage corresponding to 100% glyoxal). Effects stated at higher dosages in these two latter studies were reduced water and food intake (first study only) and retardation of body weight gain (both studies). In a study examining more sensitive end-points (serum clinical biochemistry), the lowest tested dosage of 107 mg/kg body weight per day (99% glyoxal) corresponded to the lowest-observed-adverse-effect level (LOAEL) for a 90-day exposure of rats via drinking-water. A 90-day feeding study in dogs failed to reveal any substance-related changes at the top dose of 115 mg/kg body weight per day (dose corresponding to 100% glyoxal).
In animal studies, 30% and 40% aqueous glyoxal caused slight to definite skin irritations, depending on the application time. Glyoxal is irritating to mucous membranes and acts as a skin sensitizing agent in humans and experimental animals.
Fetotoxic effects occurred only with doses of glyoxal that induced maternal toxicity. In developmental toxicity studies with rats, a NOEL for embryotoxicity was >300 mg glyoxal dihydrate/kg body weight per day (corresponding to >185 mg glyoxal/kg body weight per day), whereas a lowest-observed-effect level (LOEL) (decreased body weight gain) for maternal toxicity was 200 mg glyoxal dihydrate/kg body weight per day (corresponding to 123 mg glyoxal/kg body weight per day). Developmental toxicity range-finding studies in rabbits yielded a NOEL of 200 mg glyoxal dihydrate/kg body weight per day (corresponding to 123 mg glyoxal/ kg body weight per day) for both maternal toxicity and embryotoxicity.
Glyoxal is directly genotoxic in vitro in bacterial and mammalian cells, inducing, for example, DNA adducts, mutations, chromosomal aberrations, DNA repair, sister chromatid exchanges, and DNA single strand breaks. In vivo, a genotoxic activity of glyoxal was established at the site of application in the pyloric mucosa of rats by demonstration of unscheduled DNA synthesis and DNA single strand breaks. After oral application, DNA strand breaks were further observed in rat liver. There are no carcinogenesis bioassays with inhalation exposure to glyoxal. Glyoxal showed tumour-promoting activity in a two-stage glandular stomach carcinogenesis model in male Wistar rats, whereas it was inactive in a short-term liver foci assay. In an assay for tumour-initiating activity of glyoxal in skin and in cell transformation assays, glyoxal yielded negative test results.
Taking the 29-day inhalation study in rats exposed to glyoxal, which showed a NOEL of 0.6 mg/m3 for local effects in the larynx, and using uncertainty factors of 10 for interspecies differences and 10 for interindividual differences, a tolerable concentration of 6 µg/m3 for local effects in the larynx for short-term exposure was estimated.
In a sample risk assessment for the general population, an exposure scenario has been compiled as a hypothesized worst case. Using the daily intake of, maximally, 10 mg glyoxal via food, an estimated intake of 0.16 mg glyoxal/kg body weight per day can be calculated. This is similar to the tolerable intake of about 0.2 mg/kg body weight per day for lifetime oral exposure to glyoxal.
In a second sample risk assessment, for a nurse or hospital cleaner or consumer using disinfectant, a typical brand of disinfectant (7.5 g in 100 g = 7.5% glyoxal) is used at a dilution of 1% for disinfection and cleaning of surfaces (i.e., 0.075% glyoxal). Using a rounded-up 0.1% glyoxal solution and a calculation derived from a model gives an uptake of about 4 µg/kg body weight per day, assuming a body weight of 64 kg. This is much (50 times) less than the tolerable intake of about 0.2 mg/kg body weight per day for lifetime oral exposure. However, using a worst-case exposure to 4% glyoxal and the same assumptions as above would give an uptake of about 0.15 mg/kg body weight, which is approximately the same as the tolerable intake of about 0.2 mg/kg body weight per day for lifetime oral exposure.
In the final sample risk assessment, a farmer using a spray application of biocidal products containing glyoxal to disinfect a stable was used as an example. The model calculation using the given assumptions predicts a short-term exposure concentration of 24 µg glyoxal/m3 for a 6-min exposure and 32 µg glyoxal/m3 for a 15-min exposure. This can be compared with the estimated tolerable concentration of 6 µg/m3 for local effects in the larynx for a short-term exposure. There is a perceived risk of local laryngeal effects and irritation to the skin from this spray application of glyoxal.
Exposure to glyoxal has been shown to inhibit the activities of aerobic as well as anaerobic bacteria, green algae (96-h EC50 value of about 149 mg/litre for Pseudokirchneriella subcapitata [formerly Selenastrum capricornutum]), and an invertebrate species. In four fish species tested, the lowest reported 96-h LC50 value was 215 mg/litre (Pimephales promelas).
A sample risk characterization for the aquatic environment was performed by calculating the ratio between a local predicted environmental concentration (PEC), based on recently measured data, and a corresponding predicted no-effect concentration (PNEC). A PNEC of 149 µg/litre for surface water was estimated from the lowest EC50 value of 149 mg/litre by applying an uncertainty factor of 1000. Using the highest recently measured concentration of glyoxal in surface water (1.9 µg/litre), a PEC/PNEC quotient of 0.013 was obtained. As this is less than 1, no further information, testing, or risk reduction measures are required.
A no-observed-effect concentration (NOEC) of 68 mg/litre was determined for the inhibition of the proliferation of rhizome fragments of Helianthus tuberosus, with a corresponding EC30 value of 136 mg/litre, in the only study available. As no additional data characterizing the toxic effects exhibited by glyoxal upon terrestrial microorganisms or invertebrates are available, it was not possible to perform a reliable quantitative risk characterization.
Glyoxal (CAS No.
Glyoxal can undertake rotational isomerization between the planar cis and trans conformations, with trans-glyoxal being the more stable isomer (Bulat & Toro-Labbé, 2002):
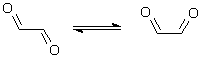
trans-glyoxal cis-glyoxal
Some of the most important hydrated derivatives of glyoxal formed by nucleophilic addition in aqueous solution are shown below (Whipple, 1970; Chastrette et al., 1983). These include the monomer ethane-1,1,2,2-tetraol (I), the dimer 2-dihydroxymethyl-(1,3)dioxolane-4,5-trans-diol (II), and the trimer bis(dioxolane) (i.e., 2,2'-bi-1,3-dioxolanyl-4,4',5,5'-tetraol) (III) — both cis and trans configurations. However, the proportion of the different structures varies with concentration and pH.
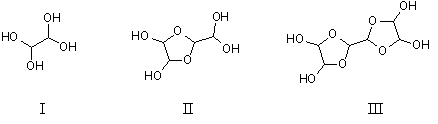
Some studies (developmental toxicity) have used glyoxal trimeric dihydrate (CAS No.
The environmentally relevant physicochemical properties of glyoxal and of the commercially employed 40% aqueous solution of this compound are summarized in Table 1. Additional physical and chemical properties are presented in the International Chemical Safety Card reproduced in this document.
The conversion factors2 for glyoxal in air (at 20 °C and 101.3 kPa) are as follows:
1 ppm in air = 2.41 mg/m3
1 mg/m3 = 0.414 ppm
Accepted methods for the detection and quantification of glyoxal in different matrices are given below. Additional and more detailed information is available in BUA (1997) and references cited therein.
Determination of glyoxal in air usually involves concentration of the alpha-dicarbonyl onto a solid sorbent coated with an appropriate derivatization agent followed by solvent desorption prior to high-performance liquid chromatographic (HPLC) detection. Zhou & Mopper (1990) reported a detection limit of about 0.05 µg/m3 for a 100-litre air sample using 2,4-dinitrophenylhydrazine (DNPH)-coated C18 cartridges, elution with carbon tetrachloride, and subsequent HPLC detection. As an alternative approach, Ho & Yu (2002) employed pentafluorobenzyl hydroxylamine (PFBHA)-coated sorbent sampling followed by thermal desorption and gas chromatography/mass spectrometry (GC/MS) detection of formed oximes and reported a minimum detection limit of 0.24 µg/m3 for a sample volume of 4.8 litres.
Table 1: Physicochemical properties of glyoxal and its commercially employed aqueous solution (40%).
|
Property |
Value |
Reference |
|
Glyoxal |
||
|
Relative molecular mass |
58.04 |
|
|
Density (g/cm3) |
1.14 (20 °C) |
Lide (1995) |
|
Refractive index |
1.3826 (20 °C) |
Lide (1995) |
|
Melting point (°C) |
15 |
Brabec (1993) |
|
Boiling point (°C) |
50.4 (101.3 kPa) |
Lide (1995) |
|
Vapour pressure (kPa) |
29.33 (~20 °C) |
Brabec (1993) |
|
n-Octanol/water partition coefficient (log Kow) |
-1.65 (calculated) |
This reporta |
|
-0.85 (measured) |
BASF AG (1988) |
|
|
Water solubility (g/litre) |
600 (25 °C) |
Hoechst AG (1994) |
|
Henry's law constant |
|
Betterton & Hoffmann (1988) |
|
(Pa·m3/mol) |
<3.38 × 10-4 (25 °C, measured) |
|
|
(dimensionless) |
<1.36 × 10-7 |
|
|
40% aqueous solution of glyoxal |
||
|
Vapour pressure (kPa) |
2.03 (20 °C) |
BASF AG (personal communication, 2003) |
|
Density (g/cm3) |
1.27 (20 °C) |
Hoechst AG (1993) |
|
Viscosity (mPa·s) |
5-10 (23 °C) |
BASF AG (1991) |
|
Setting point (°C) |
~ -10 |
Hoechst AG (1993) |
|
pH of aqueous solution |
2.1-2.7 |
Lundberg (1995) |
a Using KowWin v.1.66.
Edelkraut & Brockmann (1990) detected and quantified glyoxal in water samples by using the typical 2,4-DNPH derivatization followed by HPLC with diode array detection at 360 nm. They reported a detection limit of 295 ng/litre. Glaze et al. (1989) employed aqueous-phase PFBHA derivatization, yielding the corresponding pentafluorobenzyl oxime, followed by n-hexane extraction and detection by GC/electron capture detection (ECD) or GC/MS. A minimum detection limit of 5.1 µg/litre was obtained using GC/ECD, whereas GC/MS detection gave a minimum detection limit of 7.7 µg/litre. Method 556.1 of the US Environmental Protection Agency (US EPA, 1999) suggests a similar procedure (aqueous-phase PFBHA derivatization followed by hexane extraction and fast GC/ECD detection), leading to method detection limits in the range of 0.13-0.39 µg/litre. Steinberg & Kaplan (1984) used both HPLC and GC/MS as well as direct insertion probe/MS to detect and quantify glyoxal after 2,4-DNPH derivatization followed by dichloromethane extraction in fog and mist samples. As a viable alternative, derivatization using o-phenylenediamine to give the corresponding quinoxaline prior to HPLC/ultraviolet (UV) detection has been described (Barros et al., 1999).
As described for gaseous and liquid samples, glyoxal is usually derivatized either directly in suspended samples or after extraction by using o-phenylenediamine with subsequent GC/ECD detection or 2,4-DNPH with HPLC/UV detection. Kawata et al. (1980) found a detection limit of 0.02 mg/kg analysing sediment samples for the presence of glyoxal. No specific method for the analysis of soil is available (BUA, 1997).
The concentration of glyoxal in whole-blood samples was determined by derivatization with 1,2-diamino-4,5-dimethoxybenzene, solid-phase extraction, and HPLC of the resulting quinoxaline adduct with fluorometric detection (Thornalley et al., 1996). The interbatch coefficient of variation was 20%, the limit of detection 40 pmol, and the recovery 99%. Odani et al. (1999) employed a similar method for plasma, using quantitative derivatization of glyoxal present in plasma with 2,3-diaminonaphthalene prior to organic extraction followed by subsequent analysis employing HPLC resolution and detection by electrospray ionization/MS. Lapolla et al. (2003) quantified glyoxal in plasma using GC/MS after derivatization with O-(2,3,4,5,6-pentafluorobenzyl)hydroxylamine hydrochloride.
There are several natural sources of glyoxal. Thus, glyoxal can be produced biologically as a useful by-product (i.e., for the generation of hydrogen peroxide required by manganese-dependent peroxidase enzymes; Kersten, 1990) or non-enzymatically by autoxidation of lipids (Hirayama et al., 1984). Furthermore, it can be produced from a range of abiotic reactions with aromatic compounds in the presence of ozone and/or hydroxyl radicals. Accordingly, Mopper & Stahovec (1986) detected the formation of glyoxal from humic acids by photochemical reactions in seawater. Mopper et al. (1991) estimated the photochemical glyoxal formation rates in Sargasso seawater (0-4000 m) to be in the range of 0.4-1.1 nmol carbon/h. In addition, one can safely assume that natural fires — in analogy with results reported for domestic and residential log fires (Kleindienst et al., 1986; McDonald et al., 2000) — will release glyoxal in addition to other aldehydes. Ozone can — for example, when applied as a water disinfectant — catalyse the formation of glyoxal from organic carbon present in water (Glaze et al., 1989; Le Lacheur et al., 1991; Lopez et al., 1999).
Two well established processes employed for the production of glyoxal are the gas-phase oxidation of ethylene glycol with air in the presence of copper or silver catalysts at elevated temperature (about 300 °C) and the liquid-phase oxidation of acetaldehyde with nitric acid (Chumbhale & Awasarkar, 2001). In Germany, less than 10 000 tonnes of glyoxal (40%) were produced in 1992 (BUA, 1997). However, in 2002, BASF started up a new production plant with an annual capacity of about 60 000 tonnes (BASF AG, personal communication, 2003). The Japanese production figure for glyoxal was 13 000 tonnes in 1999 (J. Sekizawa, personal communication, 2001). The world production volume of glyoxal is about 120-170 kilotonnes (OECD, 2002).
Glyoxal is used as a chemical intermediate in the production of pharmaceuticals and dyestuffs. It is also used in the industrial production of alpha-hydroxyalkylureas (the addition of glyoxal to urea) and is industrially employed as a cross-linking agent in the production of a range of different polymers, such as textiles (e.g., permanent press fabrics) (Hoechst AG, 1984a; Choi et al., 1998, 1999; Choi, 2002), paper (Xu et al., 2002), and proteins (Marquié, 2001). It is used as a biocide and as a disinfecting agent and is present in many products, such as cleansers used for the disinfection of surfaces (BPI, 1993; OECD, 2002; BASF AG, personal communication, 2003).
Generally, glyoxal might be released during its manufacture or its application (BUA, 1997; see also section 4.3). A well recognized source of glyoxal is automotive emissions and the subsequently formed photochemical smog, which gives rise to the formation of this compound (California State Air Resources Board, 1984; Jing et al., 2001). In addition, emissions from cigarettes have been shown to contain trace amounts of glyoxal (Moree-Testa & Saint-Jalm, 1981). Another potential source of glyoxal is domestic and residential log fires (Kleindienst et al., 1986; McDonald et al., 2000). Using an irradiated smog chamber, Kleindienst et al. (1986) and McDonald et al. (2000) detected glyoxal concentrations of up to about 110 µg/m3. Interestingly, glyoxal was detected as a minor species in turbulent flames of acetylene and ethylene under atmospheric pressure (Tichy et al., 1998).
The predominant target compartments for glyoxal in the environment are the hydrosphere and soil (about 46% and 54%, respectively) and, to a lesser extent, air (<1%; Level III fugacity calculation, using EPI v.3.1). According to Thomas (1982), the reported Henry's law constant of <3.38 × 10-4 Pa·m3/mol (Betterton & Hoffmann, 1988) indicates that glyoxal is essentially non-volatile with regard to the aqueous phase. Therefore, a noteworthy transfer of glyoxal from the aqueous to the gas phase is not expected. This is supported by the findings of Harke & Höffler (1984), who used Hela cells as an indicator for the presence of inhibitory compounds (i.e., a range of biocides) in the gas phase. Their results clearly indicated that glyoxal — as opposed to other biocides tested, such as formaldehyde — was not transferred from solution to the gas phase. However, glyoxal has to be regarded as a highly mobile compound in soil due to the low log Koc value (<1) reported (BUA, 1997). Due to its excellent solubility in water and its low log Kow, it is not expected to bioaccumulate.
Atkinson (2000) calculated a lifetime of 1.1 days for glyoxal in the presence of hydroxyl radicals (assuming an average 12-h daytime concentration of 2 × 106 molecules/cm3). With respect to photolytic transformation (overhead sun), a lifetime of 5 h was calculated by the same author. Li & Schlegel (2001) showed that the photofragmentation of glyoxal proceeded — under collision-free conditions — by internal conversion to a vibrationally excited state, which dissociates to yield H2 + CO + CO (28%), H2CO (formaldehyde) + CO (65%), and HCOH (hydroxycarbene) + CO (7%). Hence, glyoxal released into the atmosphere will undergo a rapid degradation in this environmental compartment. According to Yadav & Gupta (2000), the hydrolysis of glyoxal with sodium hydroxide as catalyst proceeded with a second-order rate constant of 9.3 × 10-6 cm3/mol·s at 25 °C to yield glycolic acid. Brunet et al. (1984) showed the transformation of glyoxal to oxalic acid via glyoxylic acid in the presence of ozone.
In biodegradation tests corresponding to Organisation for Economic Co-operation and Development (OECD) guideline 301C, glyoxal was readily biodegradable (65% biochemical oxygen demand [BOD] of theoretical oxygen demand [ThOD], incubation for 14 days; MITI, 1992). Similar results were obtained by applying the method of Zahn Wellens, yielding an elimination of >70% of dissolved organic carbon in 7 days (Hoechst AG, 1991a). Conway et al. (1983) observed a significant bio-oxidation of glyoxal by sewage inocula (76% of ThOD after 20 days), while Gerike & Gode (1990) showed that glyoxal was biodegradable (i.e., 90% of ThOD within 28 days) by employing the OECD 301D closed bottle test. Furthermore, using both the oxygen consumption inhibition test and the OECD confirmatory test, they established an inhibition limit of 500 mg/litre. Hence, water or sewage treatment plants should be impacted detrimentally only at high influent concentrations. In fact, a large number of microbial enzymes catalyse the transformation of glyoxal to common intermediates in microbial catabolism. Thus, Sakai et al. (2001) reported the efficient transformation of glyoxal to glycolaldehyde by glyoxal reductase from Bacillus subtilis. Furthermore, glyoxal can be effectively oxidized by enzymes such as the fungal glyoxal oxidase (Kersten, 1990) and its bacterial counterpart (Whittaker et al., 1999) to yield glyoxylic acid, which, in turn, is a common intermediate of many metabolic sequences (i.e., glyoxylate cycle) present in microorganisms. Finally, the microbial glyoxalase system (Cooper, 1984) should yield glycolate in analogy to the reaction with methylglyoxal, whereas microbial aldehyde dehydrogenases with sufficient activity for 2-oxoaldehydes should yield glyoxylate.
Even under physiological conditions, glyoxal reacts quickly with arginine, leading to the formation of 1-(4-amino-4-carboxybutyl)-2-imino-5-oxo-imidazolidine (Schwarzenbolz et al., 1997). Thus, arginine residues present in proteins can act as scavengers for glyoxal. Further, glyoxal is able to oxidize an amino acid such as phenylalanine to a plethora of products, such as Strecker aldehydes and O- and N-heterocycles (Adamiec et al., 2001), and can produce amides from amino acids such as lysine (Glomb & Pfahler, 2001) and arginine (Glomb & Lang, 2001).
Residential wood combustion was reported to release up to about 600 mg glyoxal/kg hardwood used in the fireplace (McDonald et al., 2000). Borrego et al. (2000) detected glyoxal in ambient air sampled in the Giesta area (a rural location 20 km south-east of Aveiro, Portugal) and reported an average level of 3.7 µg/m3. Kawamura et al. (2000) showed the presence of glyoxal in all ambient air samples taken at four different locations (modest to high degree of urbanization and traffic) in the Los Angeles region (USA) and found glyoxal at concentrations ranging from about 0.096 to 2.3 µg/m3. More recently, Ho & Yu (2002) analysed the ambient air at a roadside location near a bus stop (Clear Water Bay, Kowloon) in Hong Kong for 24 h and showed that the lowest levels of glyoxal (about 1.2 µg/m3) were present in the early hours (01:00-05:00), whereas the maximum levels (about 9.9 µg/m3) were obviously correlated with increasing traffic (09:00-13:00). Jing et al. (2001) found glyoxal present in urban air samples taken in Las Vegas (USA) in both summer (range 0.29-0.99 µg/m3) and winter (range 0.22-0.51 µg/m3).
Steinberg & Kaplan (1984) detected glyoxal in fog samples collected near Los Angeles (Topanga Canyon), USA, at concentrations up to about 1.9 mg/litre.
The presence of glyoxal was reported at 4-12 µg/litre in water of the river Elbe sampled near Brunsbüttel, Germany (Edelkraut & Brockmann, 1990), and in samples from the Sargasso Sea (Mopper et al., 1991; no values given). Le Lacheur et al. (1991) detected glyoxal at low µg/litre concentrations in raw drinking-water sampled in Fort Dix, USA. More recently, Nawrocki et al. (1996) detected glyoxal in distilled (0.9 µg/litre) and double-distilled (0.1 µg/litre) reagent water. The same authors (Dabrowska et al., 2003) demonstrated the presence of glyoxal at concentrations up to 1.9 µg/litre in groundwater (Mosina water intake serving Poznañ) and surface water samples (river Bogdanka) from Poland. These levels did not significantly increase upon chlorine dioxide treatment of the raw water samples. IPCS (2000) reported the median concentration of glyoxal in ozone-treated drinking-water to be 9 µg/litre.
Kawata et al. (1980) reported the presence of glyoxal in sediment samples from Japanese rivers at up to 13 mg/kg dry weight.
Glyoxal is a substance frequently detected in fermented food and beverages. This is mainly due to microbial activity as well as non-enzymatic browning reactions such as caramelization and Maillard reactions of saccharides (Hollnagel & Kroh, 1998; Glomb & Tschirnich, 2001; Hollnagel & Kroh, 2002). Accordingly, Barros et al. (1999) found glyoxal present in different brands of beer and wine on sale in Portugal. Sampling three different brands of white wine, they detected glyoxal at concentrations of 6.2, 8.7, and 26 µmol/litre (about 360, 464, and 1509 µg/litre). De Revel & Bertrand (1993) evaluated a range of French wines and detected glyoxal in one white wine (mean of 125 µg/litre), red wines (151-368 µg/litre), and five sherry wines (lowest level of glyoxal in a Seco with 435 µg/litre and highest level in an Olorosso with 1556 µg/litre). Palamand et al. (1970) detected glyoxal levels ranging from about 230 to 1000 µg/litre in eight different beers. Nagao et al. (1986) detected glyoxal in Bourbon whiskey (390 µg/litre), wine (970 µg/litre), and apple brandy (33 µg/litre), as well as in black tea (20 µg/litre) and instant (340 µg/litre) and brewed coffee (870 µg/litre). Yamaguchi et al. (1994) detected glyoxal in beverages such as beer (20-40 µg/litre) as well as white (510 µg/litre) and red wine (740 µg/litre).
Nagao et al. (1986) found glyoxal in soybean paste (4.2 mg/kg), soy sauce (4.9 mg/litre), toast (0.5 mg/kg), and bread (0.3 mg/kg). Markianova et al. (1971) reported glyoxal levels in bread ranging from 0.07 to 0.31 mg/kg, depending on the yeast type employed. However, Roiter & Borovikova (1972) showed that using amylase in the baking process led to glyoxal levels of up to 1.4 mg/kg in the bread crust and of up to 1.6 mg/kg in the bread crumbs. Plant materials used for brewing (rice — about 14 mg/kg; barley — about 3 mg/kg; malt — about 7 mg/kg) might contain glyoxal as well (Palamand et al., 1970). Yamaguchi et al. (1994) detected glyoxal in fermented food such as yoghurt (about 0.63-0.92 mg/kg). Due to heat-induced autoxidation, edible oils might contain glyoxal, as was shown for sardine oil, containing up to 6.5 mg/kg (Hirayama et al., 1984).
The main route of exposure of the general population to glyoxal is probably via intake of water and food containing glyoxal. Glyoxal is present in a broad range of different food products. However, due to the lack of quantitative data on the presence of glyoxal in food products such as meat, dairy products, or fish, an exact value cannot be given. The general population might also be exposed to glyoxal via cigarette or residential log fire smoke or vehicle exhaust containing glyoxal.
An exposure scenario has been compiled as a hypothesized worst case. A food/drink intake of 10 mg/day has been calculated from foods with a known glyoxal content (see section 6.1.4). About three cups of brewed coffee per day (>400 µg glyoxal), toast (>50 µg glyoxal), a stir-fried meal containing rice (>4 mg glyoxal), oil (>500 µg glyoxal), soy sauce (>200 µg glyoxal), a pint of beer (500 µg glyoxal), one yoghurt (>130 µg glyoxal), and one glass of sherry (>30 µg glyoxal) leads to an intake of about 6 mg/day. A further intake of 3-4 mg glyoxal/day might come from other fermented products (dairy products or vegetables), from other popular roasted or fried products (meat, fish, mushrooms, sausages), or from additional bakery products.
Assuming a daily intake of 20 m3 air containing about 4 µg glyoxal/m3 (Borrego et al., 2000), a daily consumption of 2 litres of water containing 9 µg glyoxal/litre (median for ozone-treated drinking-water; IPCS, 2000), and an estimated daily intake of 10 mg glyoxal via food, an intake of about 160 µg of glyoxal per kg body weight (using 64 kg as the value for body weight) per day can be calculated. This intake is almost totally from food.
Modifying this calculation by including a 2-h daily exposure to traffic exhaust containing glyoxal (at 9.9 µg/m3 — from Ho & Yu (2002) — instead of 4 µg/m3) while still using the other values as stated above (water, food) does not yield a significantly higher value.
Glyoxal has been reported as being present in some household cleaners up to a concentration of 4% (product databanks, Switzerland, Denmark, and Germany; R. Hertel, personal communication, 2003). People can therefore be exposed to glyoxal during its use as a household cleaner.
Glyoxal does not appear to evaporate from solution (Harke & Höffler, 1984). Further, the reported Henry's law constant of <3.38 × 10-4 Pa·m3/mol (Betterton & Hoffmann, 1988) indicates that glyoxal is essentially non-volatile with regard to the aqueous phase. Therefore, occupational exposure by inhalation will probably take place only in situations where aerosols containing glyoxal are released. Such an exposure situation might be the spray application of biocidal products containing glyoxal.
A model calculation has been made using an aerosol droplet simulation programme for a worst-case exposure via inhalation of aerosol droplets — for example, of a farmer disinfecting his stable by spray application of a commercial product (see Appendix 5 for details). The model calculation using the given assumptions predicts an exposure concentration of 24 µg glyoxal/m3 for a 6-min exposure and 32 µg glyoxal/m3 for 15 min.
Exposure via skin (i.e., unprotected use of disinfectant solution) may be estimated using DermWin v.1.43 (US EPA, 2000). A typical brand of disinfectant (7.5 g in 100 g = 7.5% glyoxal) recommends a dilution of 1% for disinfection and cleaning of surfaces (i.e., 0.075% glyoxal). Using a rounded-up figure of 0.1% glyoxal solution and a Kp value (estimated from the following equation: log Kp = -2.72 + 0.71 log Kow — 0.0061 MW) of 5.63 × 10-5 cm/h (given in DermWin v.1.43 taken from US EPA, 2000; where Kp is the permeability coefficient from water and MW is molecular weight) for glyoxal, the dermally absorbed dose per event (assuming a final concentration of glyoxal in the aqueous solution used for cleaning surfaces of 1 mg/cm3 [0.1%] and an event duration of 30 min) yields a potential uptake of 2.8 × 10-2 µg/cm2 per event (using Fick's first law) with regard to exposed, unprotected skin. Taking a worst case of 10 events/day, a surface area of hands of 840 cm2 (US EPA, 1997), and assuming 100% uptake through the skin, this would mean 235 µg glyoxal/day, which equals 3.7 µg/kg body weight, assuming a body weight of 64 kg.
Glyoxal is produced endogenously and is commonly present in blood plasma of healthy subjects, with one study giving values of about 67 ng/ml (corresponding to about 1.16 µmol/litre; Odani et al., 1999) and other studies reporting 0.23 µmol/litre (Agalou et al., 2002) and 0.3 µmol/litre (Lapolla et al., 2003). Higher levels are found in patients with diabetes or renal diseases (see section 7.1). The urine of patients without these diseases contained glyoxal at about 132 µmol/litre (Espinosa-Mansilla et al., 1998). This value is in apparent conflict with the low levels found in tissues and body fluids and with the assumed efficient glyoxalase activities in these patients.
Glyoxal is endogenously produced during normal cellular metabolism by a multitude of enzyme-independent pathways, such as the spontaneous reaction of amino groups in proteins with reducing sugars (Maillard reaction), sugar autoxidation, DNA oxidation, peroxidation of polyunsaturated fatty acids, and UV photodamage, and in conditions of oxidative stress and depletion of GSH (Loidl-Stahlhofen & Spiteller, 1994; Shibamoto, 1994; Murata-Kamiya et al., 1995, 1997a; Wells-Knecht et al., 1995; Fu et al., 1996; Mlakar & Spiteller, 1996; Abordo et al., 1999; Miyata & Kurokawa, 1999; Thornalley et al., 1999; Kasper & Funk, 2001; Ulrich & Cerami, 2001; Kasai, 2002; Thornalley, 2002; Wondrak et al., 2002a) (see Figure 1). Furthermore, glyoxal is a product of the metabolism and microsomal oxidation of compounds such as glycolaldehyde, ethylene glycol, and beta-hydroxy-substituted N-nitrosamines and possibly contributes to the toxic, genotoxic, and tumorigenic action of these substances (Loeppky & Goelzer, 2002; Loeppky et al., 2002).
In biological materials, less than 10% of the glyoxal present is in unbound forms in aqueous solution (free glyoxal and hydrates), as most of the reactive carbonyl groups are reversibly bound to cysteinyl, lysyl, and arginyl residues of proteins (Thornalley, 1995).
The endogenous concentrations of glyoxal in human tissues and body fluids, as with other alpha-oxoaldehydes, are limited by the high catalytic efficiency of the glyoxalase system (Thornalley, 1995) as well as by the rapid reaction of glyoxal with proteins (Sady et al., 2000).
During certain pathological conditions (e.g., diabetes mellitus, uraemia), raised concentrations of glyoxal have been measured. The concentration of glyoxal in blood samples from normal human subjects (n = 19) was 0.21 ± 0.14 µmol/kg (Thornalley et al., 1996). For blood plasma, a value of approximately 0.1 µmol/litre was estimated for normal healthy subjects, which can double in diabetics (Thornalley, 1998; Thornalley et al., 2000).
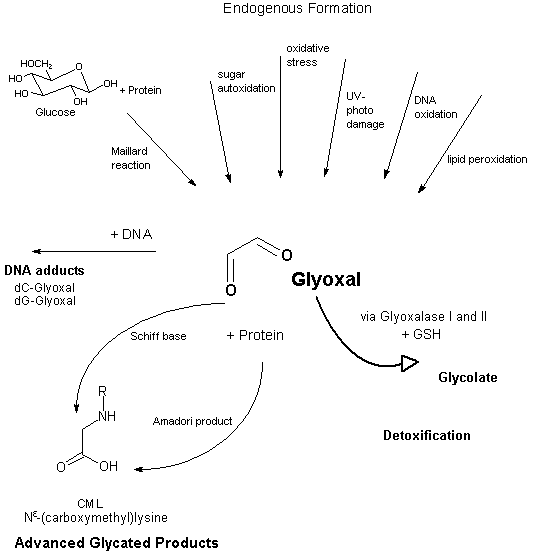
Fig. 1: Scheme of formation, detoxification, and protein and DNA adduct formation of glyoxal
The concentration of glyoxal in blood plasma was 0.23 ± 0.13 µmol/litre in controls (n = 6), 0.4 ± 0.16 µmol/litre in patients with mild/moderate uraemia (n = 10), and 0.76 ± 0.21 µmol/litre in patients with end-stage renal disease on haemodialysis (n = 5) (Agalou et al., 2002). Similar levels in plasma samples were reported by Lapolla et al. (2003) (n = 3 persons/group): a mean of 17.3 µg/litre (0.3 µmol/litre) for healthy subjects, 26.4 µg/litre (0.45 µmol/litre) for badly controlled diabetics, and 27.2 µg/litre (0.47 µmol/litre) for those affected by chronic renal failure. Another working group reported higher levels of glyoxal in plasma (n = 15-20 subjects): 67 µg/litre in normal control subjects and 78 µg/litre in patients with non-insulin-dependent diabetes, corresponding to approximately 1 µmol/litre. Chronic renal failure resulted in accumulation of glyoxal, with a mean plasma level of 221 µg/litre (about 4 µmol/litre), which was possibly caused by accelerated autoxidation of glucose in uraemic patients (Odani et al., 1999). A possible higher non-physiological production of glyoxal leading to local accumulation was assumed in patients with hyperglycaemia associated with diabetes (Akhand et al., 2001).
In porcine ischaemic heart tissue, glyoxal levels in the lipid fraction (determination of "free glyoxal") increased 4-fold after up to 4 h of ischaemia and 24-fold after 6 h of ischaemia (0.2 µg/g lipid) in comparison with extraischaemic heart tissue (Dudda et al., 1996).
In cultures of P388D1 cells (murine macrophage cell line), the intracellular background concentration was 31.2 pmol glyoxal/106 viable cells (sum of free glyoxal and glyoxal reversibly bound to proteins). Intracellularly produced glyoxal readily crosses cell membranes, possibly by passive diffusion. Despite reversible binding to cellular peptides and proteins, accumulation of glyoxal in the extracellular medium could be demonstrated, with concentrations in the culture medium rising from below the detection limit to 61 nmol/litre (P < 0.01) during a 3-h culture period (Abordo et al., 1999).
There are limited qualitative and no quantitative data on the absorption and distribution of glyoxal in humans and experimental animals. Acute and subacute inhalation exposure resulted in local effects on eyes and respiratory organs, the extent of systemic absorption being unclear. After acute and chronic oral administration, there is evidence of systemic absorption, with distribution to erythrocytes, liver, lung, kidney, pancreas, and adrenal glands (BUA, 1997; see also section 8; e.g., Ueno et al., 1991a). There is some qualitative evidence that glyoxal is absorbed after dermal exposure. Granular and vacuole degeneration in liver, kidney, and pancreas have been observed along with a distinct increase in blood glucose levels following dermal application (Ito, 1963). Further, data on skin sensitization (see sections 8.7 and 9) provide supportive qualitative evidence that glyoxal is absorbed across the skin.
In normal human urine, a glyoxal concentration of 132 µmol/litre was found by HPLC analysis (Espinosa-Mansilla et al., 1998). However, this could either be produced endogenously or stem from an exogenous source, such as food intake.
The cytosolic GSH-dependent glyoxalase system is the major pathway for the detoxification of glyoxal (see Figure 1). Glyoxal reacts non-enzymatically with GSH with formation of a hemithioacetal, which is subsequently converted to S-glycolylglutathione by glyoxalase I. Glyoxalase II catalyses the hydrolysis of S-glycolylglutathione to glycolate, re-forming the GSH from the first reaction. The activity of glyoxalase I in situ is approximately proportional to the cytosolic concentration of GSH. When GSH is severely depleted (e.g., under conditions of oxidative stress), however, 2-oxoaldehyde dehydrogenase and aldose reductase may also metabolize glyoxal. Imbalances in intracellular redox systems may impair these detoxification mechanisms, resulting in higher levels of glyoxal (Thornalley, 1995, 1998; Abordo et al., 1999; Miyata et al., 1999, 2001). A further GSH-independent route of detoxification via glyoxalase III exists. Glyoxalase III is reported to be the most abundant glyoxalase in Escherichia coli (MacLean et al., 1998; Okada-Matsumoto & Fridovich, 2000).
The glyoxalase I concentration in human tissues and blood cells was about 0.2 µg/g protein. In human tissues, the specific activity was highest in pancreas, lung, kidney, and brain and lowest in adipose tissue and liver. Specific activities in fetal tissues were about 3 times higher than in corresponding adult tissues. Human glyoxalase I was found to exhibit genetic polymorphism, with three phenotypes resulting from a diallelic gene. The frequency of the GLO1 allele in various populations on average ranges from 0.046 to 0.853 (Thornalley, 1993).
Exposure to glyoxal induced the activity of glyoxalase I in Salmonella typhimurium strains TA 100 and TA 104 (0.1 mg glyoxal/ml) (Ueno et al., 1991b) as well as in erythrocytes, liver, and kidney of male Sprague-Dawley rats (4000 or 6000 mg glyoxal/litre drinking-water for 30 days, no increased activity for longer exposure periods; for details, see section 8.3) (Ueno et al., 1991a).
Glyoxal attacks the amino groups of proteins, nucleotides, and lipids with its highly reactive carbonyl groups. A sequence of non-enzymatic reactions, called glycation, yields stable AGEs with a background extent of 0.1-1% of lysine and arginine residues in proteins and 1 in 107 nucleotides in DNA.
AGEs originating from the reaction of glyoxal with lysine and arginine residues of proteins identified so far are Nε-(carboxymethyl)lysine (CML), imidazolium cross-links as glyoxal-lysine dimer and imidazolysine, arginine-derived imidazolium products, and arginine-lysine cross-links. Cyclic imidazolidones may be formed on reaction with arginine residues.
Glyoxal forms stable adducts with guanosine by reaction with the N-1 as well as with the exocyclic nitrogen of guanine. The rate of glyoxal-guanine adduct formation is rapid under physiological conditions (Loeppky et al., 1999). A stable tricyclic glyoxal-DNA adduct is formed by covalent binding to two nitrogens of guanine under physiological conditions in vitro (for details, see BUA, 1997). Besides 8-hydroxy-deoxyguanosine, the glyoxal-deoxyguanosine (dG) adduct is one of the major deoxyguanosine oxidation products, being formed by oxygen radicals, lipid peroxidation systems, various types of oxidative stress, and UV irradiation and after in vivo exposure to beta-hydroxy-substituted N-nitrosamines (Murata-Kamiya et al., 1997a,b; Loeppky et al., 1999; Mistry et al., 1999; Cooke et al., 2000; Kasai, 2002).
Reaction of glyoxal with deoxycytidine (dC) yields 5-hydroxyacetyl-deoxycytidine or, by deamination, deoxyuridine. Deamination of 5-methyl-deoxycytidine is also possible, forming deoxythymidine. The analysis of DNA bases involved in DNA cross-links formed in vitro showed cross-linking by deoxyguanosine-glyoxal-deoxycytidine adducts and deoxyguanosine-glyoxal-deoxyadenine adducts (Kasai et al., 1998).
Incubation of rat retinal organ culture with glyoxal (<300 µmol/litre for 9 h) increased apoptotic events in all layers. After 800 µmol glyoxal/litre, approximately 50% of the cells in all layers of the retina were apoptotic. The glyoxal-induced rapid formation of CML showed the ability of the retina model to simulate AGE-related events in vitro. The neurotoxicity of glyoxal-induced AGE formation was shown by the significantly increased rate of cell death in the retina (Reber et al., 2003).
The acute toxicity of glyoxal in experimental animals is low to moderate, depending on the actual concentration of glyoxal in the tested product. However, from the documentation in the study reports, it is not always clear if the values given for the LC50 or LD50 refer to the tested product with its specified concentration or if the values were converted to a concentration of 100% glyoxal. A detailed compilation of acute toxicity data is given in the source document (BUA, 1997).
An LC50 value of 2440 mg/m3 (2410 mg/m3 for females, 2470 mg/m3 for males) was calculated from single 4-h inhalation exposures of rats to aerosols of 40% glyoxal (Hoechst AG, 1984b). All 10 rats exposed by inhalation to an atmosphere containing dust of 80% glyoxal in the highest technically feasible concentration of 1300 mg/m3 survived (Hoechst AG, 1984c). All rats survived 7- and 8-h exposures to concentrated atmospheres (concentration not further specified) of 30% (Mellon Institute, 1958, 1965) or 40% glyoxal (Hoechst AG, 1984d,e). After inhalative uptake, observations reported included local irritations of the eyes and respiratory organs as well as hyperaemia and foamy secretion in the lungs. No macroscopic organ changes were reported in those rats surviving the 14-day post-observation period (Hoechst AG, 1984d,e).
After oral administration to rats, LD50 values ranging from 2960 mg/kg body weight (lowest value in females) to 8979 mg/kg body weight (highest value in males) were reported in several studies using products containing 40% glyoxal, demonstrating a higher sensitivity of female rats. In mice (sex not given), the LD50 of 40% glyoxal was 4064 mg/kg body weight. For a preparation containing 80% glyoxal, oral LD50 values of 2000 mg/kg body weight in rats and 900 mg/kg body weight in guinea-pigs were found. Macroscopic observations reported after oral uptake include irritations of the gastrointestinal tract and congestions in the gastrointestinal tract, lung, kidney, and adrenal glands (BUA, 1997).
After dermal administration of 40% glyoxal, the LD50 values were >2000 mg/kg body weight for the rat, 12 700 mg/kg body weight for the rabbit, and >5000 mg/ kg body weight for the guinea-pig (for details, see BUA, 1997).
In the 1940s to 1960s, histopathological findings in studies with acute application of glyoxal pointed to a connection between effects induced by glyoxal and those induced in the course of diabetes; this has been confirmed by recent intensive studies on the mechanism of action of endogenous glyoxal and its involvement in the development of diabetic complications (see section 8.8).
Pancreas and kidney were identified as the prominent target organs of the toxic action of glyoxal; severe degenerative changes in these organs were attributed to an inhibition of glyoxalase activity in these tissues. Changes in the pancreas were dominated by the observation of necrotic areas containing B-cells of the Langerhans islets in rabbits (105 mg glyoxal/kg body weight intracardial or two administrations of 320 mg/kg body weight subcutaneous) and in cats (227 mg/kg body weight, application not specified). A simultaneous increase of blood glucose levels was demonstrated in rabbits and cats, comparable to alloxan-induced diabetes (Doerr et al., 1948). The pancreas is a prominent target organ of alloxan toxicity, too, which is mediated by free radicals (Younes, 1997). Rats responded to intravenous injection of 100-200 mg glyoxal/kg body weight with a dose-dependent, reversible, and reproducible reduction of the blood glucose level, which was attributed to a glyoxal-stimulated secretion of insulin secondary to oedematous changes of the pancreas. At higher dosage (175 mg/kg body weight intravenous), more severe changes, such as irreversible necroses and degranulation of B-cells, were observed in connection with visible changes in other organs. However, the B-cells of the pancreas showed the highest sensitivity to the toxic action of glyoxal (Helge, 1959). The nephrotoxic action of glyoxal is characterized by vacuole degeneration in the kidney (460 mg glyoxal per cat subcutaneous) (Doerr, 1957a,b). Acute effects noted in the pancreas in several studies all seemed to arise when glyoxal was administered parenterally, compared with other routes. This may be due to toxicokinetic reasons.
A further study in rabbits described histopathological changes in liver, kidney, and pancreas 40 days after a single dermal application of a 40% glyoxal solution (leading to severe necrotic dermatitis at application site; dose not specified). Granular and vacuole degeneration in liver, kidney, and pancreas and atrophy and fibrous change of Langerhans islets were assessed to show a close resemblance to changes in these tissues in the course of diabetes. In glucose tolerance tests performed 5 and 10 days after dermal application of glyoxal, a distinct increase of blood glucose levels was observed in comparison with a constant level in control rabbits (Ito, 1963).
In an inhalation study conducted according to OECD guideline 412, groups of five male and five female Wistar rats inhaled aerosols containing glyoxal (40% in water) at 0, 0.4, 2.0, or 10 mg/m3 (analytical concentrations 0, 0.6, 2.3, and 8.9 mg/m3; mass median aerodynamic diameter 0.8-1.2 µm) for a period of 29 days (nose only, 6 h/day, 5 days/week). Exposure was tolerated by all dose groups without any systemic effects (examination of body weight, haematological and biochemical parameters, urine analysis, macroscopic and histological examination). The only local effect found in the larynx was a minimal squamous metaplasia of the epiglottal epithelium accompanied by a minimal submucosal infiltration of lymphocytes in the mid- and high-dose groups. Consequently, for local effects, a NOEL of 0.6 mg/m3 (nominal concentration 0.4 mg/m3) resulted for subacute inhalation exposure of rats to glyoxal (Hoechst AG, 1995).
In a 28-day oral toxicity study conducted according to OECD guideline 407, six male and six female Sprague-Dawley rats per dose group were exposed to glyoxal (40% in water) at dosages of 0, 100, 300, or 1000 mg/kg body weight per day via the drinking-water. A dose-dependent retardation of body weight gain in the mid-dose group (slight effect) and the high-dose group (significant effect) was accompanied by reduced food intake. A dose-dependent reduction of water intake was observed in male rats at the lowest dose and in female rats at the mid- and high doses (glyoxal concentrations were adjusted to water intake). Changes in mid- and high-dose groups, such as increased erythrocyte number and reduced urine volume, were attributed to reduced water intake; changes of various organ weights in the high-dose group were attributed to reduced body weight. No changes were found at macroscopic and histological examination. The NOAEL for this study is 100 mg glyoxal/kg body weight per day (Société Française Hoechst, 1987). (More details were not available to the authors of this CICAD. It is not known whether these concentrations are adjusted to 100% glyoxal. If not, the NOAEL would be about 40 mg/kg body weight adjusted to 100% glyoxal.)
In a 90-day feeding study, Wistar rats (10 males and 10 females per dose group) were exposed to glyoxal (40% preparation). The study gives the dosages converted to 100% glyoxal content as corresponding to about 32, 63, 125, and 250 mg/kg body weight per day for male and female rats. Males of the high-dose group showed a reversible significant retardation of body weight gain during the first 2 weeks of exposure without a concomitant reduction of food intake. Significant increases of liver and kidney weights were observed in the high-dose group (these are the only organ weights examined). No relevant macroscopic or micropathological changes were observed in thoracic and abdominal organs (pancreas not examined). Haematological and biochemical parameters were not analysed. From these investigations, a NOAEL of 125 mg (corresponding to 100% glyoxal)/kg body weight per day was estimated (Mellon Institute, 1966).
Beagle dogs (three per dose group) were also exposed to the same preparation of glyoxal by feeding dosages of 31, 65, or 115 mg/kg body weight per day (dosages corresponding to 100% glyoxal). Up to the high dosage, no substance-related changes of body weight, food consumption, liver or kidney weight, or haematological or serum clinical chemistry parameters and no macroscopic or histopathological changes were observed in thoracic and abdominal organs (pancreas not examined). The NOEL for 90-day feeding of glyoxal to dogs was >115 mg/kg body weight per day (dosage corresponding to 100% glyoxal) (Mellon Institute, 1966).
Five male Sprague-Dawley rats per group were treated with glyoxal (98.7% purity) in drinking-water at concentrations of 2000, 4000, or 6000 mg/litre for periods of 30, 60, or 90 days (Phase I study) (Ueno et al., 1991a). Due to a decrease in food intake, the actual dosages decreased with increasing time of exposure (30, 60, and 90 days) and corresponded to 188, 135, and 107 mg/kg body weight per day for the low-dose groups, 407, 239, and 234 mg/kg body weight per day for the mid-dose groups, and 451, 344, and 315 mg/kg body weight per day for the high-dose groups, respectively. The study design included observations of clinical signs, body weights, major organ weights (liver, kidneys, spleen, heart, testes, brain), serum clinical chemistry, and biochemical examinations of glyoxalase activity and extent of lipid peroxidation (content of GSH and 2-thiobarbituric acid-reactive substances) in liver, kidneys, and erythrocytes.
There was a dose-dependent retardation of body weight gain, which was significant for the mid- and high-dose groups, and also a dose-dependent decrease of food and water intake. From Phase II of this study (see below), it was concluded that body weight reduction did not correspond to decreased food intake but was a reflection of the systemic effects of glyoxal. Absolute weight of liver, kidneys, spleen, and heart significantly decreased in all dosed groups at all time points. A significant increase of relative kidney weight in the high-dose group resulted after 90 days. There was no indication of increased lipid peroxidation.
Glyoxalase I activity was significantly increased in liver and erythrocytes at the mid- and high doses and in the kidneys at the high dose at the 30-day termination, but not for longer exposure periods. In contrast, the serum clinical parameters aspartate aminotransferase, alanine aminotransferase, lactate dehydrogenase, albumin, and total protein were significantly reduced by the mid- and/or high-dose exposures for all examination time points. In the low-dose group, alanine aminotransferase and total protein were significantly decreased, so that it was not possible to derive a NOAEL for this study. Consequently, a dosage of 107 mg/kg body weight per day (99% glyoxal) corresponds to the LOAEL for a 90-day exposure of rats (Ueno et al., 1991a). The decrease of serum protein levels was attributed to a decrease of protein synthesis, which was demonstrable after acute exposure to glyoxal (Ueno et al., 1991a) and is explainable by the mode of action of glyoxal (see section 8.8).
In Phase II of the study, five rats received 6000 mg glyoxal/litre drinking-water (highest test concentration from Phase I) for 90 or 180 days. One control group received food ad libitum, whereas a second diet-limited control group received the same amount of food as consumed by the dosed animals. Dosages were 315 and 298 mg/kg body weight per day (glyoxal 98.7% purity) for the 90- and 180-day exposure, respectively. The extent of examinations was comparable to that in Phase I and was further supplemented by gross and histopathological examinations of liver, kidneys, spleen, stomach, thymus, and mesenteric lymph nodes. Terminal body weight was significantly lower than in the pair-fed control, so that weight reduction is reflective of the systemic toxicity of glyoxal. Significant decreases of absolute weights and significant increases of relative weights of liver, kidneys, and heart were observed in glyoxal-exposed rats (Ueno et al., 1991a).
Fischer 344 rats (10 per dose group and sex) were exposed daily to drinking-water containing 0, 1000, 2000, 4000, 8000, or 16 000 mg glyoxal/litre for 90 days to establish dose ranges for a chronic study. All animals of the highest dose group were sacrificed prematurely on day 12 in a moribund state. Decreased dose-related body and organ weights as well as decreased food and water consumptions were observed at the lowest dosage. For chronic exposure, the maximum tolerated dose for rats was estimated in the range of 500-2000 mg/litre for males as the more sensitive sex (decrease of water consumption up to 28%) and 1000-4000 mg/litre for females (decrease of water consumption up to 46%) (NTP, 1991a).
In a similar study in B6C3F1 mice (10 per sex per dose group) exposed daily to drinking-water containing the same doses (0, 1000, 2000, 4000, 8000, or 16 000 mg glyoxal/litre for 90 days), all animals survived. The salient features observed were decreased body weight (decrease of 7-30% from 4000 to 16 000 mg/litre) and selected organ weights, decreased food and water consumption, and, in the male mice of all dose groups, possible chemical-related salivary gland changes (secretory depletion of submandibular gland). It was felt that the decreased water consumption (dose-dependently about 10-50%) was due to unsatisfactory palatability of the dosed water, subsequently leading to lower daily dosages and decreased feed consumption (up to 24%). From this preliminary study, recommended doses for further studies with long-term exposure were estimated to be in the range of 500-2000 mg/litre for males as the more sensitive sex (decrease of water consumption up to 12%) and 1000-4000 mg/litre for females (decrease of water consumption up to 27%) (NTP, 1991b).
No studies with long-term exposure to glyoxal by inhalation or oral routes were available.
After the exposure of Sprague-Dawley rats to dosages of 6000 mg glyoxal/litre drinking-water for up to 180 days (for details, see section 8.3), there were no neoplastic changes found at the gross and histopathological examination of liver, kidneys, spleen, stomach, thymus, and mesenteric lymph nodes (Ueno et al., 1991a).
Glyoxal showed tumour-promoting activity in a two-stage glandular stomach carcinogenesis model in male Wistar rats after an 8-week initiation treatment with N-methyl-N'-nitro-N-nitrosoguanidine in the drinking-water (100 mg/litre) along with a 10% sodium chloride dietary supplement. Subsequent promotion by exposure to glyoxal (0.5% in drinking-water from week 8 to week 40) induced significantly increased incidences of adenocarcinoma and hyperplasia in the pylorus of the glandular stomach in comparison with rats with initiation treatment only. Glyoxal treatment alone induced neither neoplastic nor hyperplastic changes in the pylorus (Takahashi et al., 1989). However, genotoxic activity (induction of unscheduled DNA synthesis and strand breaks) was demonstrated in the pyloric mucosa of the rat stomach (see section 8.5; Furihata et al., 1985, 1989; Furihata & Matsushima, 1989). A tumour-promoting potential was also derived from dose-dependent induction of ornithine decarboxylase and replicative DNA synthesis in the pyloric mucosa after a single application of 150-400 mg glyoxal/kg body weight (Furihata et al., 1985; Furihata & Matsushima, 1989, 1995).
In contrast, no tumour-promoting activity was found in a short-term liver foci assay with a 6-week glyoxal exposure via drinking-water (different concentrations of 5000 and 2000 mg/litre given in the publications) after initiation with diethylnitrosamine (single intraperitoneal dose of 200 mg/kg body weight, start of glyoxal exposure after 2-week recovery period, partial hepatectomy at week 3). Relative to the initiator-treated control group, number and area of glutathione-S-transferase placental form (GST-P) positive foci in the liver, as well as body weight, absolute liver weight, and water consumption, were significantly decreased in glyoxal-treated F344 rats (Hasegawa & Ito, 1992; Hasegawa et al., 1995).
No increase of skin tumours was observed after lifetime application of 3 µl glyoxal (two commercial products, 12.5% in water) 3 times a week to the skin of C3H/HeJ mice. Survival rates of glyoxal-treated rats were higher than those of controls. Some treated rats showed skin irritation with necrotic areas (Bushy Run, 1982).
In an assay for tumour-initiating activity, the dermal application of glyoxal alone (total initiating dose 30 mg glyoxal/mouse, 37-43% in water applied 2 times weekly for 5 weeks) induced no skin tumours in CD-1 mice within 53 weeks. After promotion by 12-O-tetradecanoyl-phorbol-13-acetate treatment for 47 weeks, 2 of 10 animals had a total of four skin papillomas, showing no significant tumour-initiating activity of glyoxal by this route (Miyakawa et al., 1991).
Glyoxal is directly genotoxic in vitro in bacterial and mammalian cells. In vivo tests show various findings. A detailed overview of genotoxicity tests in bacterial test systems is published in the source document (BUA, 1997).
In the Salmonella microsomal assay, glyoxal (test substance 30-40% glyoxal) was a direct mutagen in strains TA 100, TA 102, TA 104, and TA 2638, with a weaker response in the presence of a metabolic activation system (BUA, 1997). A direct genotoxic activity of glyoxal was further evident in the L-arabinose resistance assay with S. typhimurium BA9 and BA13 (Ruiz-Rubio et al., 1985; Ariza et al., 1988) and in the SOS chromotest with E. coli PQ37 (von der Hude et al., 1988).
Furthermore, DNA repair tests yielded positive responses in both the presence and absence of metabolic activation systems, as in the SOS umu-test with S. typhimurium TA 1535/pSK 1002 (Ono et al., 1991a,b), in the rec-assay with Bacillus subtilis (also with metabolic activation; Matsui et al., 1989), and in the differential DNA repair test with E. coli K-12/343/636 uvrB+/recA+ and K-12/343/591 uvrB-/recA- (Hellmér & Bolcsfoldi, 1992a). When the latter test was performed as a host-mediated assay in mice, with oral application of 570 or 1700 mg glyoxal/kg body weight and intravenous application of the bacteria, a genotoxic effect was not demonstrable in bacteria isolated from blood, liver, lungs, kidneys, or testicles (Hellmér & Bolcsfoldi, 1992b), which may be explained by the high reactivity of glyoxal — for example, with proteins (see section 8.8). In Saccharomyces cerevisiae D61.M, induction of mitotic recombinations pointed to reaction of glyoxal with DNA, whereas modification of proteins was indicated by chromosome losses (in the presence of propionitrile, which is a strong inducer of chromosomal malsegregation), suggesting interference of glyoxal with microtubular function (Zimmermann & Mohr, 1992).
With E. coli WP2 uvrA, in both the absence and presence of metabolic activation, negative test results were found in the standard plate incorporation assay (Hoechst AG, 1984f), whereas an insufficiently documented preincubation assay reported positive test results (Kato et al., 1989). Ueno et al. (1991b) investigated the characteristics of mutagenicity by glyoxal (particularly a possible role of active oxygen species) in S. typhimurium TA 100 and TA 104. The scavengers of singlet oxygen almost completely suppressed the mutagenic action of glyoxal.
A direct genotoxic action of glyoxal was established in a variety of tests with mammalian cells without metabolic activation (see BUA, 1997): in a mutagenicity test with mouse lymphoma cells (TK assay) (Wangenheim & Bolcsfoldi, 1988), in chromosomal aberration tests with Chinese hamster ovary (CHO) cells (NOTOX, 1986) and V79 cells (Nishi et al., 1989), and in tests for the induction of unscheduled DNA synthesis in TC-SV40 cells of Syrian hamster (Cornago et al., 1989), for the induction of sister chromatid exchanges in CHO cells and human lymphocytes, for the induction of endoreduplication in CHO cells (Tucker et al., 1989), and for the induction of DNA strand breaks in mouse lymphoma cells (Garberg et al., 1988). In primary rat hepatocytes, glyoxal induced DNA single strand breaks but no DNA cross-links (Ueno et al., 1991c).
DNA damage was further demonstrated in the comet assay with TK6 human lymphoblastoid cells by the induction of concentration-dependent increases of tail moment and tail length (Henderson et al., 1998). Primary rat hepatocytes exposed to glyoxal at higher concentrations (0.5-10 mg/ml) produced different concentration-dependent types of DNA damage. Tail moment and the formation of comets with head and tail (indicative of DNA strand breakage) decreased with increasing glyoxal concentration, whereas circular DNA spots with highly condensed areas increasingly appeared at the mid- and high concentrations. Among 100 tested substances, this damage was shown to be specific for certain aldehydes and was attributed to their DNA cross-linking activity (Kuchenmeister et al., 1998). In cultures of human umbilical vein endothelial cells, addition of 100 µg glyoxal/ml caused a significant increase of formamidopyrimidine N-glycosylase (FPG)-sensitive sites (measured by the comet assay) in the absence of increased intracellular levels of hydroperoxides. FPG repairs oxidative DNA damage and abasic sites and further was supposed to repair guanine-glyoxal adducts (Shimoi et al., 2001).
A significantly increased rate of sex-linked recessive lethals reported in Drosophila melanogaster in preliminary experiments (Mazar Barnett & Muñoz, 1969) was not confirmed in later assays, showing the absence of any genotoxic effect in assays for sex-linked recessive lethals in mature sperm and in the earlier stages of spermatogenesis, as well as in assays for clastogenic activity in mature sperm (reciprocal translocation, dominant lethal, and chromosome loss). However, from the increase of radiation-induced clastogenic effects after pretreatment with glyoxal, it was concluded that glyoxal came in contact with the target cells. The possibility of detoxifying mechanisms for glyoxal or of an efficient repair of glyoxal-induced damage in Drosophila was discussed (Mazar Barnett & Muñoz, 1989).
No clastogenic activity was found in a micronucleus assay in mouse bone marrow (Société Française Hoechst, 1986; no further data available).
Glyoxal was demonstrated to be genotoxic at the site of application after administration by gastric intubation. In the pyloric mucosa of male Fischer 344 rats, both significantly increased unscheduled DNA synthesis and DNA single strand breaks were induced at dosages of 400-500 mg/kg body weight within 2 h. Cytotoxicity was not reported (Furihata et al., 1985, 1988, 1989; Furihata & Matsushima, 1989). In contrast, in rat hepatocytes, a test for unscheduled DNA synthesis was negative (CCR, 1992). Glyoxal has also been shown to cause DNA strand breaks in rat hepatocytes 2-9 h after a single oral exposure to 200-1000 mg glyoxal/kg body weight (Ueno et al., 1991b). Single strand breaks were also detected in livers of rats within 2 h following a single oral exposure at 200-1000 mg glyoxal/kg body weight. The frequency of breaks reached a maximum after 9 h of exposure. Hardly any DNA lesions were detected in other tissues following exposure to 1000 mg glyoxal/kg body weight. Glyoxal causes DNA single strand breaks in rat hepatocytes following in vitro and in vivo exposure (Ueno et al., 1991c).
Cell transformation assays in C3H/10T½ cells with three different commercial products of glyoxal (test concentrations from 0.0013 to 0.195 µl/ml) yielded negative test results (Mason 1980a,b,c).
There were no studies available on the effects of glyoxal on fertility.
In developmental toxicity studies with Sprague-Dawley rats, glyoxal trimeric dihydrate was administered by gavage on gestation days 6-15 at doses of 0, 200, 800, 1200, 1600, or 2000 mg/kg body weight per day in the preliminary study and 50, 150, or 300 mg/kg body weight per day in the main study (NTP, 1991c, 1994a,b). In the initial range-finding study, maternal toxicity (decreased weight gain) was observed at 200 mg glyoxal dihydrate/kg body weight per day (corresponding to 123 mg glyoxal/kg body weight per day), with clinical signs of toxicity and decreased gravid uterine weight at 800 mg glyoxal dihydrate/kg body weight per day and maternal deaths at 1200 mg/kg body weight per day and above (NTP, 1991c). No maternal toxicity was observed, however, in the second study at the top dose of 300 mg/kg body weight per day (corresponding to 185 mg glyoxal/kg body weight per day). No embryotoxicity was observed at 200 mg glyoxal dihydrate/kg body weight per day in the preliminary study or at the highest dose in the main study.
In a study conducted according to OECD guideline 414, 40% glyoxal was administered to 19-24 female Wistar rats as a solution in water at doses of 0, 5, 25, or 125 mg/kg body weight per day (related to 100% glyoxal) on days 6 through 19 post-coitum. Maternal toxicity (significantly reduced food consumption and significantly lower corrected body weight gain) was observed at 125 mg/kg body weight per day. No substance-related effects were observed on gestational parameters or fetuses. NOAELs of 25 mg/kg body weight per day for maternal toxicity and 125 mg/kg body weight per day for embryotoxicity were established (BASF & Clariant, 2000).
Developmental toxicity range-finding studies in New Zealand White rabbits administered glyoxal by gavage yielded a NOEL of 200 mg glyoxal trimeric dihydrate/kg body weight per day, corresponding to 123 mg glyoxal/kg body weight per day (NTP, 1991d), and a LOEL of 400 mg glyoxal dihydrate/kg body weight per day, corresponding to 247 mg glyoxal/kg body weight per day (NTP, 1992), for both maternal toxicity and embryotoxicity. Maternal signs of systemic toxicity and decreases of weight parameters were accompanied by reduced fetal weight (NTP, 1992). The application of doses in the range of 200 mg glyoxal dihydrate/kg body weight per day was discussed as problematic due to the corrosive nature of the substance, leading to damage of gastric mucosa of pregnant rabbits (unpublished observations cited in NTP, 1993). In a subsequent study with a single dose level of 50 mg glyoxal dihydrate/kg body weight per day, corresponding to 31 mg glyoxal/kg body weight per day, there was no maternal mortality or persistent signs of toxicity, although minimal reductions in body weight gain and food consumption were noted. Glyoxal exposure did not significantly alter post-implantation loss and had no effect on fetal body weight or the incidence of external, visceral, or skeletal malformations. The authors gave a NOAEL for developmental toxicity for rabbits of 50 mg glyoxal dihydrate/kg body weight per day, corresponding to 31 mg glyoxal/kg body weight per day (NTP, 1993).
In an investigation on the effects of ethylene glycol and its metabolites, glyoxal was tested in a whole rat embryo culture test system (head-fold rat embryos, which lack biotransforming enzyme activities, such as alcohol dehydrogenase or acetaldehyde dehydrogenase) (Klug et al., 2001). At glyoxal concentrations of 3 mmol/ litre, the rate of dysmorphogenic embryos was within historical controls; at 6 mmol/litre, however, a general embryotoxic effect of glyoxal was noted, with a steep concentration-response relationship. At 10 mmol/litre, no growth or differentiation of the embryo could be determined. The relevance of this test for the in vivo toxicity of glyoxal is unclear.
After a 4-h exposure patch test on rabbits (OECD guideline 404), glyoxal (40%) caused no irritation. However, in earlier studies (e.g., patch test on the shaven back skin of white rabbits using 30% or 40% glyoxal), slight to pronounced irritation could be seen, depending on the application period (1 min, 5 min, 15 min, and 20 h). A single dermal administration (occlusive) of 40% glyoxal to the shaven skin (dorsal, dorsolateral) of five Wistar rats per sex for 24 h caused erythemas in all animals (BUA, 1997).
A 40% glyoxal solution was applied to the shaven back skin of white rabbits (no details of the time of administration). From the 3rd day, a strong reddened inflammation was observed, followed by a necrosis with tissue demarcation. Histopathological examination showed severe necrotic skin changes on the 4th day and a regeneration of the epidermis on the 18th day (Ito, 1963).
Therefore, taking into account studies where the duration of exposure was longer, glyoxal is clearly irritating to the skin (see details in BUA, 1997).
In a study conducted according to OECD guideline 405, glyoxal (40%) led to a reversible reddening and chemosis of the conjunctiva within 8 days and thus showed an irritating effect. This confirmed older studies reporting that glyoxal caused irritations and even necrotic changes in the rabbit eye (see details in BUA, 1997).
Two studies using the Magnusson and Kligman test and one study using the Buhler test on guinea-pigs gave positive results. Glyoxal can be considered as a sensitizing substance (see details in BUA, 1997).
Glyoxal has been shown to be sensitizing in humans (see section 9).
Glyoxal, which attacks amino groups of proteins, nucleotides, and lipids, is considered an important intermediate in the formation of AGEs. AGE modification alters protein function and inactivates enzymes, resulting in disturbance of cellular metabolism, impaired proteolysis, and inhibition of cell proliferation and protein synthesis (Gan & Ansari, 1986; Ueno et al., 1991a,b; Kasper et al., 1999; Witowski et al., 2000; Bulteau et al., 2001; Kasper & Funk, 2001; Murata-Kamiya & Kamiya, 2001). The extent of AGE modification increases with the increasing life span of proteins. Consequently, AGEs are especially associated with long-lived proteins, such as collagens, lens crystallins, and neurofilaments, but also have been identified in shorter-lived proteins, including haemoglobin, plasma proteins, lipoproteins, and intracellular proteins. AGEs have a role in the pathogenesis or progression of many pathological conditions — e.g., diabetes, Alzheimer's disease and other neurodegenerative diseases, chronic inflammatory diseases, arthritis, atherosclerosis, vascular damage, cataract formation and skin changes during ageing, pulmonary fibrosis, and renal failure — as well as in peritoneal dialysis complications (Miyata et al., 1999, 2000; Thornalley et al., 1999; Cerami & Ulrich, 2001; Ulrich & Cerami, 2001; Thornalley, 2002). Although glyoxal is known to be an intermediate in the formation of AGEs, these effects have not specifically been shown for glyoxal.
Inhibition studies in bacterial mutagenicity tests demonstrated the production of the reactive oxygen species superoxide, hydrogen peroxide, and singlet oxygen from glyoxal. The mutagenic activity of glyoxal is related to singlet oxygen, as well as to the intracellular GSH level (Suwa et al., 1982; Garst et al., 1983; Yamaguchi & Nakagawa, 1983; Ueno et al., 1991b). The hydroxyl radical plays a prominent role in glyoxal-induced DNA cleavage (Roberts et al., 2003).
The sensitizing potential of glyoxal is attributed to the electron-deficient alpha,beta-dicarbonyl structure with its high electrophilic reactivity. Glyoxal easily forms Schiff bases with lysine or arginine units and so becomes bound to skin proteins (Roberts et al., 1999).
Only limited information on the effects of glyoxal in humans has been identified.
The oral ingestion of 50-300 ml of a disinfectant containing 7.5% (w/w) glyoxal, 9.5% glutaral, and 9.6% didecyldimethylammoniumchloride in suicidal intention caused severe coagulative necrosis of the mucous membranes in the upper gastrointestinal tract and whole respiratory tract in a 31-year-old female. Despite the severe effects, no perforation occurred, probably because mucous membranes were denatured and fixed through the carbonyl groups of the aldehydes, so that the substances did not penetrate deeply (Menzner et al., 1998).
Of 14 workers who had contact with 40% glyoxal, 9 exhibited a contact dermatitis with localizations mainly on the lower arms and fingers. Patch tests with a 20% glyoxal solution produced a positive reaction in 7 of 9 workers (Ito, 1963).
In a German multicentre study of dermal sensitivity, the records of 31 849 health care workers from 24 allergy departments between 1992 and 1995 were evaluated; 4.2% of the 774 female patients working in the medical profession were found to show positive reactions to glyoxal patch testing, whereas only 1.4% of the control group (1895 persons not in the medical profession) were found to be positive (Schnuch et al., 1998).
In a continuation of this multicentre study, between 1997 and 1999, 2689 patients were reported to have been patch tested with glyoxal (trimer; 1% in petrolatum). Positive (allergic) reactions were observed in 1.6% of the patients, whereas irritant (0.3%) and questionable (0.6%) (i.e., non-allergic) reactions were observed less frequently. Even prior to diagnostic patch testing, disinfectants had been suspected to be causative for dermatitis in 23 and cleaning agents in 10 of the 44 patients sensitized to glyoxal. Occupations included nursing (n = 15), room cleaning (n = 12), dental nursing (n = 5), geriatric nursing (n = 3), and some other medical associated professions (n = 5), with very few other non-medical occupations (n = 4) (Uter et al., 2001).
In a retrospective descriptive analysis of records from an occupational dermatitis clinic in Osnabrück, Germany, 189 patients with occupational dermatitis were patch tested with glyoxal (15 in water or, as trimer, 1% in petrolatum) between 1993 and 1999. Of the 11 cases with positive reactions to glyoxal, 9 were considered as being occupationally relevant — those with nursing and room cleaning jobs (Uter et al., 2001).
In another study of 280 health care workers in Poland, the frequency of allergy to aldehydes (1% formaldehyde, glutaraldehyde, or glyoxal) with allergic dermatitis was 22.8%. The majority were sensitive to only one aldehyde, indicating lack of cross-reaction; glutaraldehyde was positive in 12.4% and glyoxal in 1.9% (Kieć-Świerczyńska et al., 1998). In a further study of the causes of occupational dermatosis in 27 dental nurses during the years 1995-1999, contact sensitization by glyoxal was found in 3 cases (Kieć-Świerczyńska & Kręcisz, 2000).
In a maximization test, all 24 patients tested gave a positive reaction with 10% glyoxal solution (induction), 2% solution (challenge) showing a very strong skin sensitizing potential (Kligman, 1966).
Table 2: Aquatic and terrestrial toxicity of glyoxal.
|
Species tested |
End-point |
Concentration |
Reference |
|
Bacteria |
|
|
|
|
Pseudomonas putida |
16-h EC10 |
46 |
Hoechst AG (1989) |
|
Pseudomonas putida |
EC0 |
500 |
Gerike & Gode (1990) |
|
Photobacterium phosphoreum |
5-min EC50 |
755 ± 55 |
Chou & Que Hee (1992) |
|
Anaerobes (not characterized) |
24-h EC0 |
200 |
Hoechst AG (1984f) |
|
Algae |
|
|
|
|
Pseudokirchneriella subcapitata (formerly Selenastrum capricornutum) |
96-h EC50 |
149 |
Bollman et al. (1990) |
|
Invertebrates |
|
|
|
|
Daphnia magna (water flea) |
24-h EC50 |
430 |
OECD (1992) |
|
Vertebrates |
|
|
|
|
Brachydanio rerio (zebra danio) |
24-h LC50 |
1200 |
Hoechst AG (1991b) |
|
Leuciscus idus melanotus (orfe) |
48-h LC50 |
>680 |
BUA (1997) |
|
Pimephales promelas (fathead minnow) |
24-h LC50 |
550 |
Conway et al. (1983) |
|
Rhombus maximus (turbot) |
48-h EC50 |
>500 |
Hoechst AG (1990) |
|
Plants |
|
|
|
|
Helianthus tuberosus (Jerusalem artichoke) |
EC30 |
136 |
BUA (1997) |
Glyoxal may enter the hydrosphere due to its production, use, and application and can be formed, in addition, by natural processes.
A limited number of acute tests have been performed to establish the toxicity of glyoxal for aquatic organisms representing different trophic levels (data are summarized in Table 2; see BUA, 1997, for more detailed information). Assays performed by employing aerobic heterotrophic bacteria such as Pseudomonas putida, anaerobic bacteria, and Photobacterium phosphoreum showed that P. putida (end-point = inhibition of cell proliferation, 16 h) exhibited the lowest EC50 value of 134 mg/litre (Hoechst AG, 1989).
The only algal species tested was Pseudokirchneriella subcapitata (formerly Selenastrum capricornutum), with a 96-h EC50 value of 149 mg/litre (Bollman et al., 1990).
For the only invertebrate species tested, Daphnia magna, a 24-h EC50 value of 430 mg/litre was reported (OECD, 1992).
Acute toxicity studies conducted with four different fish species established the lowest LC50 value (96-h incubation) as 215 mg glyoxal/litre using Pimephales promelas (Conway et al., 1983).
Data concerning the toxicity of glyoxal for soil-bound microbial activity, terrestrial invertebrates, or vertebrates or effects upon terrestrial ecosystems are not available. For the terrestrial compartment, the only available toxicity study reported the inhibition of rhizome fragment proliferation of Helianthus tuberosus by glyoxal, with a NOEC value of 68 mg/litre and a corresponding EC30 value of 136 mg/litre (BUA, 1997).
Glyoxal is endogenously produced during normal cellular metabolism by several enzyme-independent pathways. The cytosolic GSH-dependent glyoxalase system is the major pathway for the detoxification of glyoxal. When GSH is severely depleted, 2-oxoaldehyde dehydrogenase and aldose reductase also metabolize glyoxal.
Due to its highly reactive carbonyl groups, glyoxal attacks proteins, nucleotides, and lipids, followed by further reactions leading to the formation of AGEs. These adducts can interfere with normal cellular function, inducing carbonyl stress and oxidative stress and affecting protein function and signal transduction pathways of the cells; these result in a range of pathological changes, cell proliferation, genotoxicity, or programmed cell death.
During certain pathological conditions (e.g., diabetes mellitus, uraemia), raised concentrations of glyoxal have been measured in the plasma.
It is not known if acute environmental exposure to glyoxal also leads to raised concentrations in the blood or whether the high catalytic efficiency of the glyoxalase system is able to detoxify it.
Studies in patients and volunteers have confirmed the sensitizing potential of glyoxal. This has been substantiated by animal studies. Glyoxal is irritating to mucous membranes. In animal studies, 30% and 40% aqueous glyoxal cause slight to definite skin irritations, depending on the application time.
There are almost no data on other toxicity end-points in humans.
Because of the limited nature of data in humans, hazard identification and dose-response analysis for glyoxal are based primarily on studies in animals.
The acute toxicity of glyoxal in experimental animals is low to moderate. After exposure by inhalation, local irritations of the eyes and respiratory organs predominate. After oral uptake of glyoxal, macroscopic observations include irritation of the gastrointestinal tract and congestion in the gastrointestinal tract, lung, kidney, and adrenal glands. In the pancreas and kidney, the toxic action of glyoxal leads to severe degenerative changes resembling those induced during diabetes.
A 29-day nose-only inhalation exposure study in rats using 40% glyoxal showed a NOEL of 0.6 mg/m3 (nominal concentration was 0.4 mg/m3) for local effects in the larynx and a NOEL of >8.9 mg/m3 (nominal concentration was 10 mg/m3) for systemic effects (Hoechst AG, 1995).
The 90-day feeding study in rats resulted in a NOAEL of 125 mg/kg body weight per day (dose corresponding to 100% glyoxal). A 28-day drinking-water study of 40% glyoxal in rats resulted in a NOAEL of 100 mg glyoxal/kg body weight per day (Société Française Hoechst, 1987). Effects at higher doses were reduced water and food intake and retardation of body weight gain (Mellon Institute, 1966). A 90-day feeding study in dogs failed to reveal any substance-related changes at the top dose of 115 mg/kg body weight per day (dose corresponding to 100% glyoxal) (Mellon Institute, 1966).
In a 90-day drinking-water study in rats examining more sensitive end-points, the lowest tested dosage of 107 mg/kg body weight per day (99% glyoxal) was given as the LOAEL for serum clinical parameters (Ueno et al., 1991a).
There are no data available on the effects of glyoxal on fertility. Fetotoxic and developmental effects occur only with doses of glyoxal that induce maternal toxicity.
Glyoxal is directly genotoxic in vitro in bacterial and mammalian cells. In vivo, a genotoxic activity of glyoxal was established at the site of application in the pyloric mucosa of rats by demonstration of unscheduled DNA synthesis and DNA single strand breaks. After oral application, DNA strand breaks were further observed in rat liver. Glyoxal forms stable adducts with proteins and DNA bases.
No carcinogenicity studies were available for exposure by inhalation. Glyoxal showed tumour-promoting activity in a two-stage glandular stomach carcinogenesis model in male Wistar rats (Takahashi et al., 1989), whereas it was inactive in a short-term liver foci assay (Hasegawa & Ito, 1992; Hasegawa et al., 1995). In a skin painting study for tumour-initiating activity and in cell transformation assays, glyoxal yielded negative test results. A lifetime skin painting study showed no increase of tumours, but some treated rats showed skin irritation with necrotic areas.
Due to lack of data, it is not possible to determine whether glyoxal has a carcinogenic potential. It is, however, genotoxic in vitro in bacterial and mammalian cells, and there is some evidence that this may be so in vivo. It readily forms DNA adducts, generating potential carcinogens such as glyoxalated deoxyguanosine and deoxycytidine.
Exposure to exogenous glyoxal causes local effects, probably due to formation of AGEs. Occupational exposure would be mainly due to the use of glyoxal in disinfectants and adhesives and would be via inhalation of aerosols or dermal routes, causing irritant or sensitizing effects.
A 29-day inhalation study in rats exposed to glyoxal showed a NOEL of 0.6 mg/m3 for local effects in the larynx. Use of uncertainty factors of 10 for interspecies differences and 10 for interindividual differences gives a tolerable concentration of 6 µg/m3 for local effects in the larynx for short-term exposure.
From studies on oral exposure, the NOAEL seems to be about 100 mg/kg body weight per day (adjusted to 100% glyoxal). Using uncertainty factors of 10 for interspecies differences and 10 for interindividual differences and a factor of 5 for less-than-lifetime exposure, this results in a tolerable intake of about 0.2 mg/kg body weight per day for lifetime oral exposure to glyoxal. The short- and medium-term studies seem to have similar outcomes, with no evidence of systemic effects, suggesting that exogenous glyoxal is efficiently detoxified and does not accumulate in the body. The use of the lifetime extrapolation uncertainty factor (factor of 5) is also justified on the basis of the 125 mg/kg body weight LOAEL with wide dose spacing to a NOAEL of 25 mg/kg body weight (BASF & Clariant, 2000).
Example 1 — General population: An exposure scenario has been compiled as a hypothesized worst case. Using the daily intake of, maximally, 10 mg glyoxal via food given in section 6.2.1, an estimated intake of 0.16 mg glyoxal/kg body weight per day can be calculated. This is slightly less than the tolerable intake of about 0.2 mg/kg body weight per day for lifetime oral exposure to glyoxal (see section 11.1.2).
Example 2 — A nurse or hospital cleaner or consumer using disinfectant: A typical brand of disinfectant (7.5 g in 100 g = 7.5% glyoxal) is used at a dilution of 1% for disinfection and cleaning of surfaces (i.e., 0.075% glyoxal). Using a rounded-up 0.1% glyoxal solution and a calculation derived from a model gives an uptake of about 4 µg/kg body weight per day, assuming a body weight of 64 kg (see section 6.2.2).
This is much (50 times) less than the tolerable intake of about 0.2 mg/kg body weight per day for lifetime oral exposure (see section 11.1.2).
However, it should be noted that other substances (e.g., glutaral or formaldehyde) may also be present in the product.
An exposure scenario has been compiled as a hypothesized worst case. Assuming exposure to 4% glyoxal given in section 6.2.1 and using the same assumptions as above would give an uptake of about 0.15 mg/kg body weight, which is slightly less than the tolerable intake of about 0.2 mg/kg body weight per day for lifetime oral exposure (see section 11.1.2).
However, it should be noted that dermal contact to glyoxal may cause sensitization.
Example 3 — A farmer using a spray application of biocidal products containing glyoxal to disinfect a stable (see section 6 and Appendix 5): The model calculation using the given assumptions predicts a short-term exposure concentration of 24 µg glyoxal/m3 for a 6-min exposure and 32 µg glyoxal/m3 for 15 min. This can be compared with the estimated tolerable concentration of 6 µg/m3 for local effects in the larynx for a short-term exposure (see section 11.1.2). There is a perceived risk of local laryngeal effects and irritation to the skin from this spray application of glyoxal.
Glyoxal is produced endogenously during normal cellular metabolism. Glyoxal attacks proteins, nucleotides, and lipids, followed by further reactions leading to the formation of AGEs. It is uncertain as to the effects of exogenously administered glyoxal. It is possible that the detoxification mechanisms (e.g., cytosolic GSH-dependent glyoxalase system) are sufficient to counteract this. There are, however, no data available to confirm this.
There are no data on the effects of glyoxal in humans, except for sensitization effects.
There is a lack of data on the carcinogenicity of glyoxal, in particular via inhalation and oral routes.
There is little known about the toxicokinetics of glyoxal.
There is little known about the ability of stable protein adducts to accumulate in long-life cells (e.g., retinal neurons), even at normal plasma and tissue concentrations.
There is no information on occupational exposure in the glyoxal production industry.
There are no dermal exposure data for, for example, hospital staff frequently in contact with this chemical.
There are uncertainties with regard to model calculations.
Glyoxal is present in products together with other chemicals. Therefore, the risk assessment given here is for glyoxal, but not for the product itself.
The main environmental target compartments of glyoxal are soil and water. Glyoxal is quickly transformed by abiotic reactions and readily biodegraded. At present, a reliable quantification of glyoxal currently released from all sources is impossible with the data available.
The reported low log Kow values along with the high solubility in water indicate a negligible bioaccumulation potential for glyoxal.
The biocidal activity of the dialdehyde glyoxal is related to the availability of the carbonyl group. However, due to the hydration of glyoxal in the presence of water (by means of a nucleophilic addition), this activity is apparently reduced, as indicated by inhibition studies with bacteria (Eggensperger, 1977).
A sample risk characterization may be performed for glyoxal present in the hydrosphere, according to EC (1996), by calculating the ratio between a PEC (based on measured data) and a corresponding PNEC.
Due to the limited data available, a reliable quantification of glyoxal released into the environment is not possible. However, up-to-date monitoring data from Poland may be regarded as pertinent for an industrialized region. The highest reported value for surface water samples of the river Bogdanka of 1.9 µg/litre can be employed as a local PEC. A corresponding PNEC for surface water can be predicted from the lowest 96-h EC50 value obtained for Pseudokirchneriella subcapitata growth inhibition (149 mg/litre) using an uncertainty factor of 1000 (EC, 1996). Thus, PNEC = 149 mg/litre / 1000 = 0.149 mg/litre.
Employing the highest recently measured concentration of glyoxal in surface water, the PEC/PNEC ratio (1.9 µg/litre / 149 µg/litre) gives a risk quotient of 0.013. As this value is clearly smaller than 1, no further information, testing, or risk reduction measures are required. Further, using the maximum but older value measured in surface water, 12 µg/litre in the river Elbe, the risk quotient PEC/PNEC is still less than 1.
Glyoxal will preferably partition into soil and water (about 54% and 46%, respectively; Level III fugacity calculation) and only to a lesser extent into air.
For the terrestrial compartment, the only available toxicity study measured the inhibition of rhizome fragment proliferation of Helianthus tuberosus by glyoxal (NOEC = 68 mg/litre).
The reported log Koc values indicate a high mobility in soil and point to a potential to leach into groundwater. However, the ready biodegradability, rapid abiotic transformation, and negligible bioaccumulation potential indicate a low tendency of glyoxal to pose a risk to the terrestrial compartment. As there is a lack of valid data concerning the ecotoxicity of this simple dialdehyde for relevant soil indicator organisms, a risk characterization is not possible.
The acute toxicity of glyoxal was tested using several aquatic species from different trophic levels. However, the amount of data is still limited; in particular, long-term toxicity studies with invertebrates or vertebrates are not available.
No toxicity studies are available for sediment-dwelling organisms.
For the terrestrial compartment, the only available toxicity study measuring the inhibition of rhizome fragment proliferation of Helianthus tuberosus by glyoxal appears not to be sufficient to support a quantitative risk characterization.
There are no previous evaluations of glyoxal by international bodies.
Abordo EA, Minhas HS, Thornalley PJ (1999) Accumulation of alpha-oxoaldehydes during oxidative stress: a role in cytotoxicity. Biochemical Pharmacology, 58:641-648.
Adamiec J, Rössner J, Velisek J, Cejpek K, Savel J (2001) Minor Strecker degradation products of phenylalanine and phenylglycine. European Food Research and Technology, 212:135-140.
Agalou S, Karachalias N, Thornalley PJ, Tucker B, Dawnay AB (2002) Estimation of alpha-oxoaldehydes formed from the degradation of glycolytic intermediates and glucose fragmentation in blood plasma of human subjects with uraemia. Excerpta Medica International Congress Series, 1245:181-182.
Akhand AA, Hossain K, Kato M, Miyata T, Du J, Suzuki H, Kurokawa K, Nakashima I (2001) Glyoxal and methylglyoxal induce aggregation and inactivation of ERK in human endothelial cells. Free Radical Biology and Medicine, 31(10):1228-1235.
Ariza RR, Dorado G, Barbancho M, Pueyo C (1988) Study of the cause of direct-acting mutagenicity in coffee and tea using the Ara test in Salmonella typhimurium. Mutation Research, 201:89-96.
Atkinson R (2000) Atmospheric chemistry of VOCs and NOx. Atmospheric Environment, 34(12-14):2063-2101.
Barros A, Rodrigues JA, Almeida PJ, Oliva-Teles MT (1999) Determination of glyoxal, methylglyoxal, and diacetyl in selected beer and wine, by HPLC with UV spectrophotometric detection, after derivatization with o-phenylenediamine. Journal of Liquid Chromatography and Related Technology, 22(13):2061-2069.
BASF AG (1988) 1-Octanol-Wasser. Verteilungskoeffizient von Glyoxal bei 25°C. Unpublished report, 16 August (BRU 88.157) [cited in BUA, 1997].
BASF AG (1991) DIN-Sicherheitsblatt Glyoxal P. Ludwigshafen, BASF AG, June.
BASF AG, Clariant (2000) Glyoxal 40% — prenatal development toxicity study in Wistar rats. Unpublished report (BASF No. 30R0146/99011) [cited in OECD, 2002].
Betterton EA, Hoffmann MR (1988) Henry's law constants of some environmentally important aldehydes. Environmental Science and Technology, 22:1415-1418.
Bollman MA, Banne WK, Smith S, DeWhitt K, Kapustka L (1990) Report on algal toxicity tests on selected Office of Toxic Substances (OTS) chemicals. Corvallis, OR, US Environmental Protection Agency, pp. 1-42 (EPA/600/3-90/041; PB-90-212606).
Borrego C, Gomes P, Barros N, Miranda AI (2000) Importance of handling organic atmospheric pollutants for assessing air quality. Journal of Chromatography A, 889(1-2):271-279.
BPI (1993) Rote Liste 1993, Pharmaceutical list of the Bundesverband der Pharmazeutischen Industrie e. V. (BPI). Aulendorf/Württenburg, Editio Cantor.
Brabec MJ (1993) Aldehydes and acetals. In: Clayton GD, Clayton FE, eds. Patty's industrial hygiene and toxicology, 4th ed., Vol. 2A. New York, NY, John Wiley & Sons, pp. 283-327.
Brunet R, Bourbigot MM, Dore M (1984) Oxidation of organic compounds through the combination ozone-hydrogen peroxide. Ozone Science & Engineering, 6:163-183.
BUA (1997) [Glyoxal.] German Chemical Society (GDCh) Advisory Committee on Existing Chemicals of Environmental Relevance (BUA). Stuttgart, S. Hirzel, Wissenschaftliche Verlagsgesellschaft, pp. 1-64 (BUA Report 187) (in German).
Bulat F, Toro-Labbé A (2002) A theoretical study of the rotational isomerization of glyoxal and halogen derivatives. Chemical Physics Letters, 354:508-517.
Bulteau A-L, Verbekes P, Petropoulos I, Chaffotte A-F, Friguet B (2001) Proteasome inhibition in glyoxal-treated fibroblasts and resistance of glycated glucose-6-phosphate dehydrogenase to 20 S proteasome degradation in vitro. Journal of Biological Chemistry, 276(49):45662-45668.
Bushy Run (1982) Evaluation of the dermal carcinogenicity of Aerotex glyoxal 40 and European glyoxal 40 in male C3H mice. Prepared by Bushy Run Research Center, Export, PA, for the American Cyanamide Company (Unpublished Report No. 45-508) [cited in BUA, 1997].
California State Air Resources Board (1984) Formation and fate of toxic chemicals in California's atmosphere. Sacramento, CA, California State Air Resources Board, July, 38 pp. (PB85-172609).
CCR (1992) In vivo/in vitro unscheduled DNA synthesis in rat hepatocytes with glyoxal. Prepared by Cytotest Cell Research GmbH & Co. KG, Roßdorf, for the Employment Accident Insurance Fund of the German Chemical Industry (Unpublished Report CCR Project 230602) [cited in BUA, 1997].
Cerami A, Ulrich P (2001) Pharmaceutical intervention of advanced glycation endproducts. Novartis Foundation Symposium, 235:202-220.
Chastrette F, Bracoud C, Chastrette M, Mattioda G, Christidis Y (1983) Etude de la composition de solutions aqueuses de glyoxal en RMN-13C. Bulletin de la Société Chimique de France, 2:33-40.
Choi HM (2002) Effects of boron compounds on cotton fabrics treated with glyoxal for non-formaldehyde wrinkle resistant finishing. American Association of Textile Chemists and Colorists Review, 2:42-45.
Choi HM, Park MJ, Oh KW (1998) Nonformaldehyde crease-resistant finishing of silk with glyoxal. Textile Chemist and Colorist, 30(12):41-45.
Choi HM, Kim JH, Shin S (1999) Characterization of cotton fabrics treated with glyoxal and glutaraldehyde. Journal of Applied Polymer Science, 73(13):2691-2699.
Chou CC, Que Hee SS (1992) Microtox EC50 values for drinking water by-products produced by ozonolysis. Ecotoxicology and Environmental Safety, 23(3):355-363.
Chumbhale VR, Awasarkar PA (2001) Oxidative dehydrogenation of ethylene glycol into glyoxal over phosphorus-doped ferric molybdate catalyst. Applied Catalysis, 205:109-115.
Conway RA, Waggy GT, Spiegel MH, Berglund RL (1983) Environmental fate and effects of ethylene oxide. Environmental Science and Technology, 17(2):107-112.
Cooke MS, Mistry N, Ladapo A, Herbert KE, Lunec J (2000) Immunochemical quantitation of UV-induced oxidative and dimeric DNA damage to human keratinocytes. Free Radical Research, 33(4):369-381.
Cooper RA (1984) Metabolism of methylglyoxal in microorganisms. Annual Review of Microbiology, 38:49-68.
Cornago P, Lopez Zumel M, Santos L, Pintado M (1989) Semiconservative and unscheduled DNA synthesis on mammalian cells and its modification by glyoxylic compounds. Biochimie, 71:1205-1210.
Dabrowska A, Swietlik J, Nawrocki J (2003) Formation of aldehydes upon ClO2 disinfection. Water Research, 37:1161-1169.
de Revel G, Bertrand A (1993) A method for the detection of carbonyl compounds in wine: glyoxal and methylglyoxal. Journal of the Science of Food and Agriculture, 61:267-272.
Doerr W (1957a) Über Entzündung und Degeneration. Deutsche Medizinische Wochenschrift, 82:685-691.
Doerr W (1957b) Über Entzündung und Degeneration. Deutsche Medizinische Wochenschrift, 82:713-714.
Doerr W, Bopp F, Kuhn R, Quadbeck G (1948) Pankreasschäden durch Glyoxal. Naturwissenschaften, 4:125-127.
Dudda A, Spiteller G, Kobelt F (1996) Lipid oxidation products in ischemic porcine heart tissue. Chemistry and Physics of Lipids, 82(1):39-51.
EC (1996) Technical guidance document in support of the Commission Directive 93/EEC on risk assessment for new notified substances and the Commission Regulation (EC)1488/94 on risk assessment for existing substances. Ispra, European Commission, European Chemicals Bureau.
Edelkraut F, Brockmann U (1990) Simultaneous determination of carboxylic acids and carbonyl compounds in estuaries by HPLC. Chromatographia, 30:432-435.
Eggensperger H (1977) Zur antimikrobiellen Wirkung von Aldehyden. Hospital-Hygiene, Gesundheitswesen und Desinfektion, 3:79-85.
Espinosa-Mansilla A, Durán-Merás I, Salinas F (1998) High-performance liquid chromatographic-fluorometric determination of glyoxal, methylglyoxal, and diacetyl in urine by prederivatization to pteridinic rings. Analytical Biochemistry, 255:263-273.
Fu M-X, Requena JR, Jenkins AJ, Lyons TJ, Baynes JW, Thorpe SR (1996) The advanced glycation end product, Nε-(carboxymethyl)lysine, is a product of both lipid peroxidation and glycoxidation reactions. Journal of Biological Chemistry, 271:9982-9986.
Furihata C, Matsushima T (1989) Prediction of possible carcinogens, tumor-promotors and anti-tumor promotors in the glandular stomach. Environmental and Molecular Mutagenesis, 14(15):63.
Furihata C, Matsushima T (1995) In vivo short-term assays for tumor initiation and promotion in the glandular stomach of Fischer rats. Mutation Research, 339:15-35.
Furihata C, Yoshida S, Matsushima T (1985) Potential initiating and promoting activities of diacetyl and glyoxal in rat stomach mucosa. Japanese Journal of Cancer Research, 76(9):809-814.
Furihata C, Sato Y, Matsushima T (1988) Alkaline elution of DNA from stomach pyloric mucosa of rats treated with MNNG and glyoxal. Mutation Research, 203:371.
Furihata C, Hatta A, Sato Y, Matsushima T (1989) Alkaline elution of DNA from stomach pyloric mucosa of rats treated with glyoxal. Mutation Research, 213:227-231.
Gan JC, Ansari GAS (1986) Non-oxidative inactivation of plasma alpha1-proteinase inhibitor by carbonyl compounds found in cigarette smoke. Research Communications in Substances of Abuse, 7(1-2):59-69.
Garberg P, Akerblom EL, Bolcsfoldi G (1988) Evaluation of a genotoxicity test measuring DNA-strand breaks in mouse lymphoma cells by alkaline unwinding and hydroxyapatite elution. Mutation Research 203:155-176.
Garst J, Stapleton P, Johnston J (1983) Mutagenicity of alpha-hydroxy ketones may involve superoxide anion radical. In: Greenwald RA, Cohen G, eds. Oxy radicals and their scavenger system. Vol. 2. Cellular and medical aspects. New York, NY, Elsevier, pp. 125-130.
Gerike P, Gode P (1990) The biodegradability and inhibitory threshold concentration of some disinfectants. Chemosphere, 21(6):799-812.
Glaze WH, Koga M, Cancilla D (1989) Ozonation byproducts. 2. Improvement of an aqueous-phase derivatization method for the detection of formaldehyde and other carbonyl compounds formed by the ozonation of drinking water. Environmental Science and Technology, 23:838-847.
Glomb MA, Lang G (2001) Isolation and characterization of glyoxal-arginine modifications. Journal of Agricultural and Food Chemistry, 49:1493-1501.
Glomb MA, Pfahler C (2001) Amides are novel protein modifications formed by physiological sugars. Journal of Biological Chemistry, 276(45):41638-41647.
Glomb MA, Tschirnich R (2001) Detection of alpha-dicarbonyl compounds in Maillard reaction systems and in vivo. Journal of Agricultural and Food Chemistry, 49(11):5543-5550.
Harke HP, Höffler J (1984) Übergang antimikrobieller Wirkstoffe von der Fläche in die Luft. Hygiene und Medizin, 9:259-260.
Hasegawa R, Ito N (1992) Liver medium-term bioassay in rats for screening of carcinogens and modifying factors in hepatocarcinogenesis. Food and Chemical Toxicology, 30(11):979-992.
Hasegawa R, Ogiso T, Imaida K, Shirai T, Ito N (1995) Analysis of the potential carcinogenicity of coffee and its related compounds in a medium-term liver bioassay of rats. Food and Chemical Toxicology, 33(1):15-20.
Helge H (1959) Über den Glyoxaleffekt auf Pankreas und Blutzucker. Verhandlungen der Deutschen Gesellschaft für Pathologie, 42:158-163.
Hellmér L, Bolcsfoldi G (1992a) An evaluation of the E. coli K-12 uvrB/recA DNA repair host-mediated assay. I. In vitro sensitivity of the bacteria to 61 compounds. Mutation Research, 272:145-160.
Hellmér L, Bolcsfoldi G (1992b) An evaluation of the E. coli K-12 uvrB/recA DNA repair host-mediated assay. II. In vivo results for 36 compounds tested in the mouse. Mutation Research, 272:161-173.
Henderson L, Wolfreys A, Fedyk J, Bourner C, Windebank S (1998) The ability of the comet assay to discriminate between genotoxins and cytotoxins. Mutagenesis, 13(1):89-94.
Hirayama T, Yamada N, Nohara M, Fukui S (1984) The existence of the 1,2-dicarbonyl compounds glyoxal, methyl glyoxal and diacetyl in autoxidised edible oils. Journal of the Science of Food and Agriculture, 35:1357-1362.
Ho SSH, Yu JZ (2002) Feasibility of collection and analysis of airborne carbonyls by on-sorbent derivatization and thermal desorption. Analytical Chemistry, 74:1232-1240.
Hoechst AG (1984a) Glyoxal. Broschüre der Société Française Hoechst. Paris, Hoechst AG, pp. 1-32.
Hoechst AG (1984b) Glyoxal 40N. Akute Aerosolinhalation an männlichen und weiblichen SPF-Wistar-Ratten. 4 Stunden — LC50. Frankfurt, Hoechst AG, pp. 1-7 (Report No. 84.0378; NTIS/OTS 0535127; EPA/OTS 86-920000799).
Hoechst AG (1984c) Glyoxal 80. Akute Staubinhalation an männlichen und weiblichen SPF-Wistar-Ratten. 4 Stunden — LC50. Frankfurt, Hoechst AG, pp. 1-11 (Report No. 84.0693; NTIS/OTS 0535128; EPA/OTS 86-920000800).
Hoechst AG (1984d) Glyoxal 40N. Inhalationstoxizität im Zeitsättigungstest an männlichen und weiblichen SPF-Wistar-Ratten. Frankfurt, Hoechst AG, 24 July, pp. 8-15 (Unpublished Report No. 84.0443).
Hoechst AG (1984e) Glyoxal 40T. Inhalationstoxizität im Zeitsättigungstest an männlichen und weiblichen SPF-Wistar-Ratten. Frankfurt, Hoechst AG, pp. 1-7 (Report No. 84.0450; NTIS/OTS 0535126; EPA/OTS 86-920000798).
Hoechst AG (1984f) Ergebnis der abwasserbiologischen Untersuchungen: Glyoxal 40. Frankfurt, Hoechst AG, pp. 1-4 (W 84-087).
Hoechst AG (1989) Untersuchung auf Bakterienschädlichkeit: Zellvermehrungs-Hemmtest. Frankfurt, Hoechst AG, pp. 1-2 (V 89-74-B).
Hoechst AG (1990) Glyoxal: 96-hour acute toxicity study in turbot (Rhombus maximus). Frankfurt, Hoechst AG, 30 May, pp. 1-19 (Report No. 90.0486).
Hoechst AG (1991a) Bericht über die biologische Abbaubarkeit von Glyoxal 40 % T im Zahn-Wellens-Test gemäß vorliegender Laboraufzeichnungen von 1984. Frankfurt, Hoechst AG, 20 December, pp. 1-6 (84-0105-W1).
Hoechst AG (1991b) Personal communication of the Société Française Hoechst, September, 11 pp. [cited in BUA, 1997].
Hoechst AG (1993) DIN-Sicherheitsdatenblatt Glyoxal 40. Frankfurt, Hoechst AG, Marketing Tenside und Hilfsmittel, pp. 1-2.
Hoechst AG (1994) IUCLID data sheet on glyoxal [cited in BUA, 1997].
Hoechst AG (1995) Glyoxal 40% (aqueous solution) — testing for subacute (29-days) inhalation toxicity in male and female Wistar rats. Frankfurt, Hoechst AG, Pharma Development, Corporate Toxicology (Unpublished Report No. 94.1056) [cited in BUA, 1997].
Hollnagel A, Kroh LW (1998) Formation of alpha-dicarbonyl fragments from mono- and disaccharides under caramelization and Maillard reaction conditions. Zeitschrift für Lebensmittel-Untersuchung und -Forschung A, 207:50-54.
Hollnagel A, Kroh LW (2002) 3-Deoxypentosulose: An alpha-dicarbonyl compound predominating in nonenzymatic browning of oligosaccharides in aqueous solution. Journal of Agricultural and Food Chemistry, 50(6):1659-1664.
IPCS (2000) Disinfectants and disinfectant by-products. Geneva, World Health Organization, International Programme on Chemical Safety, 499 pp. (Environmental Health Criteria 216).
IPCS (2002) Glyoxal (40% solution). Geneva, World Health Organization, International Programme on Chemical Safety (International Chemical Safety Card 1162).
Ito K (1963) Glyoxal as a cause of occupational disease. Bulletin of the Pharmaceutical Research Institute (Osaka), 44:8-15.
Jing LH, Steinberg SM, Johnson BJ (2001) Aldehyde and monocyclic aromatic hydrocarbon mixing ratios at an urban site in Las Vegas, Nevada. Journal of the Air and Waste Management Association, 51(9):1359-1366.
Kasai H (2002) Chemistry-based studies on oxidative DNA damage: formation, repair, and mutagenesis. Free Radical Biology and Medicine, 33(4):450-456.
Kasai H, Iwamoto-Tanaka N, Fukada S (1998) DNA modifications by the mutagen glyoxal: adduction to G and C, deamination of C and GC and GA cross-linking. Carcinogenesis, 19(8):1459-1465.
Kasper M, Funk RHW (2001) Age-related changes in cells and tissues due to advanced glycation end products (AGEs). Archives of Gerontology and Geriatrics, 32(3):233-243.
Kasper M, Schinzel R, Niwa T, Munch G, Witt M, Fehrenbach H, Wilsch-Bräuninger M, Pehlke K, Hofer A, Funk RHW (1999) Experimental induction of AGEs in fetal L132 lung cells changes the level of intracellular cathepsin. Biochemical and Biophysical Research Communications, 261:175-182.
Kato F, Araki A, Nozaki K, Matsushima T (1989) Mutagenicity of aldehydes and diketones. Mutation Research, 216:366-367.
Kawamura K, Steinberg S, Kaplan IR (2000) Homologous series of C1-C10 monocarboxylic acids and C1-C6 carbonyls in Los Angeles air and motor vehicle exhausts. Atmospheric Environment, 34(24):4175-4191.
Kawata K, Ozaki K, Mukai H (1980) Gas chromatographic determination of micro amount of glyoxal in water and sediment. Bunseki Kagaku, 29:517-522.
Kersten PJ (1990) Glyoxal oxidase of Phanerochaete chrysosporium: its characterization and activation by lignin peroxidase. Proceedings of the National Academy of Sciences of the United States of America, 87:2936-2940.
Kieć-Świerczynśka M, Kręcisz B (2000) [Causes of occupational allergy in dental nurses. An analysis based on the material collected at the Institute of Occupational Medicine in Lodz.] Medycyna Pracy, 51(2):145-149 (in Polish).
Kieć-Świerczynśka M, Kręcisz B, Krysiak B, Kuchowicz E, Rydzynski K (1998) Occupational allergy to aldehydes in health care workers. Clinical observations. Experiments. International Journal of Occupational Medicine and Environmental Health, 11:349-358.
Kleindienst T, Shepson P, Edney E (1986) Wood smoke: Measurement of the mutagenic activities of its gas- and particulate-phase photooxidation products. Environmental Science and Technology, 20:493-501.
Kligman A (1966) The identification of contact allergens by human assay. III. The maximization test: a procedure for screening and rating contact sensitizers. Journal of Investigative Dermatology, 47(5):393-409.
Klug S, Merker HJ, Jäckh R (2001) Effects of ethylene glycol and metabolites on in vitro development of rat embryos during organogenesis. Toxicology in Vitro, 15(6):635-642.
Kuchenmeister F, Schmezer P, Engelhardt G (1998) Genotoxic bifunctional aldehydes produce specific images in the comet assay. Mutation Research, 419(1-3):69-78.
Lapolla A, Flamini R, Tonus T, Fedele D, Senesi A, Reitano R, Marotta E, Pace G, Seraglia R, Traldi P (2003) An effective derivatization method for quantitative determination of glyoxal and methylglyoxal in plasma samples by gas chromatography/mass spectrometry. Rapid Communications in Mass Spectrometry, 17(8):876-878.
Le Lacheur R, Singer P, Charles M (1991) Disinfection by-products in New Jersey drinking waters. Journal of the American Water Works Association, 61(5):791-805.
Li X, Schlegel HB (2001) Photodissociation of glyoxal: Resolution of a paradox. Journal of Chemical Physics, 114:8-10.
Lide DR (1995) CRC handbook of chemistry and physics, 76th ed. Boca Raton, FL, CRC Press.
Loeppky RN, Goelzer P (2002) Microsome-mediated oxidation of N-nitrosodiethanolamine (NDELA), a bident carcinogen. Chemical Research in Toxicology, 15(4):457-469.
Loeppky RN, Cui W, Goelzer P, Park M, Ye Q (1999) Glyoxal-guanine DNA adducts: detection, stability and formation in vivo from nitrosamines. IARC Scientific Publications, 150:155-168.
Loeppky RN, Ye Q, Goelzer P, Chen Y (2002) DNA adducts from N-nitrosodiethanolamine and related beta-oxidized nitrosamines in vivo: 32P-postlabeling methods for glyoxal- and O6-hydroxyethyldeoxyguanosine adducts. Chemical Research in Toxicology, 15:470-482.
Loidl-Stahlhofen A, Spiteller G (1994) alpha-Hydroxyaldehydes, products of lipid peroxidation. Biochimica et Biophysica Acta, 1211(2):156-160.
Lopez A, Ricco G, Ciannarella R, Rozzi A, Di Pinto AC, Passino R (1999) Textile wastewater reuse: Ozonation of membrane concentrated secondary effluent. Water Science and Technology, 40(4-5):99-105.
Lundberg P (1995) Glyoxal. The Nordic Expert Group for Criteria Documentation of Health Risks from Chemicals. Arbete Och Hälsa, 2(116):1-17.
MacLean MJ, Ness LS, Ferguson GP, Booth IR (1998) The role of glyoxalase I in the detoxification of methylglyoxal and in the activation of the KefB K+ efflux system in Escherichia coli. Molecular Microbiology, 27(3):563-571.
Markianova LM, Borovikova LA, Permilowskaya SW (1971) Action of yeast hybrids on the level of carbonyl compounds in bread. Khlebopekarnaya i Konditerskaya Promyshiennost, 15:18-19.
Marquié C (2001) Chemical reactions in cottonseed protein cross-linking by formaldehyde, glutaraldehyde, and glyoxal for the formation of protein films with enhanced mechanical properties. Journal of Agricultural and Food Chemistry, 49(10):4676-4681.
Mason (1980a) C3H/ 10T1/ 2 cell transformation assay, Aerotex glyoxal 40. Prepared by EG & G Mason Research Institute, Rockville, MD, for the American Cyanamide Company (Unpublished Report No. 029-626-292-8) [cited in BUA, 1997].
Mason (1980b) C3H/ 10T1/ 2 cell transformation assay, European glyoxal 40. Prepared by EG & G Mason Research Institute, Rockville, MD, for the American Cyanamide Company (Unpublished Report No. 029-626-293-8) [cited in BUA, 1997].
Mason (1980c) C3H/ 10T1/ 2 cell transformation assay, American Hoechst Glyoxal 40. Prepared by EG & G Mason Research Institute, Rockville, MD, for the American Cyanamide Company (Unpublished Report No. 029-636-321-8) [cited in BUA, 1997].
Matsui S, Yamamoto R, Yamada H (1989) The Bacillus subtilis/ microsome rec-assay for the detection of DNA damaging substances which may occur in chlorinated and ozonated waters. Water Science and Technology, 21:875-887.
Mazar Barnett B, Muñoz ER (1969) Mutation test with glyoxal in Drosophila melanogaster males. Drosophila Information Service, 44:119.
Mazar Barnett B, Muñoz ER (1989) Effect of glyoxal pretreatment on radiation-induced genetic damage in Drosophila melanogaster. Mutation Research, 212:173-179.
McDonald JD, Zielinska B, Fujita EM, Sagebiel JC, Chow JC, Watson JG (2000) Fine particle and gaseous emission rates from residential wood combustion. Environmental Science and Technology, 34(11):2080-2091.
Mellon Institute (1958) Special report on range finding tests on eight purity glyoxal 29.2% solution. Pittsburgh, PA, University of Pittsburgh, Mellon Institute of Industrial Research, 12 August, pp. 1-3 (Report 21-74; NTIS/OTS 953-5072).
Mellon Institute (1965) Special report. Range finding tests on glyoxal, 40%. Pittsburgh, PA, University of Pittsburgh, Mellon Institute of Industrial Research, 2 June, pp. 1-4 (Report 26-103; NTIS/OTS 953-5072).
Mellon Institute (1966) Special report. Results of feeding glyoxal in the diet of rats and of dogs for three months. Pittsburgh, PA, University of Pittsburgh, Mellon Institute of Industrial Research, 3 January, pp. 1-20 (Report 29-1; NTIS/OTS 852-5072).
Menzner A, Ferber A, Weilemann LS (1998) Suizidversuch mit einem klinischen Desinfektionsmittel. Intensivmedizin und Notfallmedizin, 35(4):281-285.
Mistry N, Evans MD, Griffiths HR, Kasai H, Herbert KE, Lunec J (1999) Immunochemical detection of glyoxal DNA damage. Free Radical Biology and Medicine, 26(9/10):1267-1273.
MITI (1992) Biodegradation and bioaccumulation: Data of existing chemicals based on the CSCL Japan. Tokyo, Ministry of International Trade & Industry, October, pp. 1-27, 2-63.
Miyakawa Y, Nishi Y, Kato K, Sato H, Takahasi M, Hayashi Y (1991) Initiating activity of eight pyrolysates of carbohydrates in a two stage mouse skin tumorigenesis model. Carcinogenesis, 12(7):1169-1179.
Miyata T, Kurokawa K (1999) Carbonyl stress: increased carbonyl modification of proteins by autoxidation products of carbohydrates and lipids in uremia. International Journal of Artificial Organs, 22:195-198.
Miyata T, van Ypersele de Strihou C, Kurokawa K, Baynes JW (1999) Alterations in nonenzymatic biochemistry in uremia: origin and significance of "carbonyl stress" in long-term uremic complications. Kidney International, 55:389-399.
Miyata T, Kurokawa K, van Ypersele de Strihou C (2000) Relevance of oxidative and carbonyl stress to long-term uremic complications. Kidney International, 76:120-125.
Miyata T, van Ypersele de Strihou C, Imasawa T, Yoshino A, Ueda Y, Ogura H, Kominami K, Onogi H, Inagi R, Nangaku M, Kurokawa K (2001) Glyoxalase I deficiency is associated with an unusual level of advanced glycation end product in a hemodialysis patient. Kidney International, 60(6):2351-2359.
Mlakar A, Spiteller G (1996) Previously unknown aldehydic lipid peroxidation compounds of arachidonic acid. Chemistry and Physics of Lipids, 79(1):47-53.
Mopper K, Stahovec WL (1986) Sources and sinks of low molecular weight organic carbonyl compounds in seawater. Marine Chemistry, 19:305-321.
Mopper K, Zhou X, Kieber R, Kieber D, Sikorski R, Jones R (1991) Photochemical degradation of dissolved organic carbon and its impact on the oceanic carbon cycle. Nature, 353:60-62.
Moree-Testa P, Saint-Jalm Y (1981) Determination of alpha-dicarbonyl compounds in cigarette smoke. Journal of Chromatography, 217:197-218.
Murata-Kamiya N, Kamiya H (2001) Methylglyoxal, an endogenous aldehyde, crosslinks DNA polymerase and the substrate DNA. Nucleic Acids Research, 29(16):3433-3438.
Murata-Kamiya N, Kamiya H, Iwamoto N, Kasai H (1995) Formation of a mutagen, glyoxal, from DNA treated with oxygen free radicals. Carcinogenesis, 16:2251-2253.
Murata-Kamiya N, Kamiya H, Muraoka M, Kaji H, Kasai H (1997a) Comparison of oxidation products from DNA components by gamma-irradiation and Fenton-type reactions. Journal of Radiation Research, 38(2):121-131.
Murata-Kamiya N, Kamiya H, Kaji H, Kasai H (1997b) Glyoxal, a major product of DNA oxidation, induces mutations at G:C sites on a shuttle vector plasmid replicated in mammalian cells. Nucleic Acids Research, 25(10):1897-1902.
Nagao M, Fujita Y, Wakabayashi K, Nukaya H, Kosuge T, Sugimura T (1986) Mutagens in coffee and other beverages. Environmental Health Perspectives, 67:89-91.
Nawrocki J, Kalkowska I, Dabrowska A (1996) Optimization of solid-phase extraction method for analysis of low-ppb amounts of aldehydes — ozonation by-products. Journal of Chromatography A, 749:157-163.
Nishi Y, Miyakawa Y, Kato K (1989) Chromosome aberrations induced by pyrolysates of carbohydrates in Chinese hamster V79 cells. Mutation Research, 227:117-123.
NOTOX (1986) Evaluation of the ability of SIS 503 to induce chromosome aberrations in cultured Chinese hamster ovary (CHO) cells. Prepared by NOTOX C.V., 's-Hertogenbosch, for Henkel KGaA, Düsseldorf (Unpublished Report No. 0367/EC 124) [cited in BUA, 1997].
NTP (1991a) A subchronic toxicity report of glyoxal by dosed water in Fischer-344 rats. Research Triangle Park, NC, National Institutes of Health, National Toxicology Program, 12 June, pp. 1-3 (SRI-Chm-91-523; NO1-ES-05289).
NTP (1991b) A subchronic toxicity report of glyoxal by dosed water in B6C3F1 mice. Research Triangle Park, NC, National Institutes of Health, National Toxicology Program, 14 June, pp. 1-3 (SRI-Chm-91-534; NO1-ES-05289).
NTP (1991c) Range finding studies: Developmental toxicity, glyoxal trimeric dihydrate when administered via gavage to CD Sprague-Dawley rats. Research Triangle Park, NC, National Institutes of Health, National Toxicology Program (Study No. NTP-90-RF/DT-014; NIEHS/NTP Contract No. NO1-ES-95249).
NTP (1991d) Range finding studies: Developmental toxicity, glyoxal dihydrate when administered via gavage in New Zealand White rabbits. Research Triangle Park, NC, National Institutes of Health, National Toxicology Program, December, pp. 1-14 (NTP-91-RF/DT-022).
NTP (1992) Range finding studies: Developmental toxicity, glyoxal dihydrate (repeat) when administered via gavage in New Zealand White rabbits. Research Triangle Park, NC, National Institutes of Health, National Toxicology Program, June, pp. 1-23 (NTP-92-RF/DT-030).
NTP (1993) Final report on the developmental toxicity of glyoxal trimeric dihydrate (CAS No.
NTP (1994a) Final report on the developmental toxicity of glyoxal trimeric dihydrate (CAS #
NTP (1994b) Final report on the developmental toxicity of glyoxal trimeric dihydrate (CAS #
Odani H, Shinzato T, Matsumoto Y, Usami J, Maeda K (1999) Increase in three alpha,beta-dicarbonyl compound levels in human uremic plasma: specific in vivo determination of intermediates in advanced Maillard reaction. Biochemical and Biophysical Research Communications, 256(1):89-93.
OECD (1992) Glyoxal, CAS
OECD (2002) OECD integrated HPV database. Paris, Organisation for Economic Co-operation and Development. Available at http://cs3-hq.oecd.org/scripts/hpv.
Okada-Matsumoto A, Fridovich I (2000) The role of alpha,beta-dicarbonyl compounds in the toxicity of short chain sugars. Journal of Biological Chemistry, 275:34853-34857.
Ono Y, Somiya I, Kawamura M (1991a) The evaluation of genotoxicity using DNA repairing test for chemicals produced in chlorination and ozonation process. Water Science and Technology, 23:329-338.
Ono Y, Somiya I, Kawamura M (1991b) Genotoxicity of by-products in the chemical oxidation processes. Water Science and Technology, 14:633-641.
Palamand S, Nelson G, Hardwick W (1970) Further studies on glyoxal and methylglyoxal in beer. In: Proceedings of the Annual Meeting of the American Society of Brewing Chemists. St. Paul, MN, American Society of Brewing Chemists, pp. 186-191.
Reber F, Geffarth R, Kasper M, Reichenbach A, Schleicher DE, Siegner A, Funk WHR (2003) Graded sensitiveness of the various retinal neuron populations on the glyoxal-mediated formation of advanced glycation end products and ways of protection. Graefe's Archive for Clinical and Experimental Ophthalmology, 241:213-225.
Roberts DW, York M, Basketter DA (1999) Structure-activity relationships in the murine local lymph node assay for skin sensitization: alpha,beta-diketones. Contact Dermatitis, 41(1):14-17.
Roberts MJ, Wondrak GT, Laurean DC, Jacobson MK, Jacobson EL (2003) DNA damage by carbonyl stress in human skin cells. Mutation Research, 522(1-2):45-56.
Roiter IM, Borovikova LA (1972) Level of volatile carbonyl compounds in bread during the addition of enzyme preparations. Khlebopekarnaya i Konditerskaya Promyshiennost, 14:14-15.
Ruiz-Rubio M, Alejandro-Duran E, Pueyo C (1985) Oxidative mutagens specific for A.T. base pairs induce forward mutations to L-arabinose resistance in Salmonella typhimurium. Mutation Research, 147:153-163.
Sady C, Jiang CL, Chellan P, Madhun Z, Duve Y, Glomb MA, Nagaraj RH (2000) Maillard reactions by alpha-oxoaldehydes: detection of glyoxal-modified proteins. Biochimica et Biophysica Acta, 1481:255-264.
Sakai A, Katayama K, Katsuragi T, Tani Y (2001) Glycoaldehyde-forming route in Bacillus subtilis in relation to vitamin B6 biosynthesis. Journal of Bioscience and Bioengineering, 91(2):147-152.
Schnuch A, Uter W, Geier J, Frosch PJ, Rustemeyer T (1998) Contact allergies in healthcare workers. Results from the IVDK. Acta Dermato-Venereologica, 78:358-363.
Schwarzenbolz U, Henle T, Klostermeyer RHH (1997) On the reaction of glyoxal with proteins. Zeitschrift für Lebensmittel-Untersuchung und -Forschung, 205:121-124.
Shibamoto T (1994) The role of lipid peroxidation caused by ultraviolet light in skin diseases. Journal of Toxicology — Cutaneous and Ocular Toxicology, 13(3):193-202.
Shimoi K, Okitsu A, Green MHL, Lower JE, Ohta T, Kaji K, Terato H, Ide H, Kinae N (2001) Oxidate DNA damage by high glucose and its suppression in human umbilical vein endothelial cells. Mutation Research, 480:371-378.
Société Française Hoechst (1986) Glyoxal 40% N. In vivo mutagenicity study, micronucleus test in mice. Unpublished report (Study No. 2018 MAS; HOE 86.0783) [cited in BUA, 1997].
Société Française Hoechst (1987) Glyoxal 40. 28 day dose range finding study in rats by administration in drinking water. Unpublished report (Study No. 2619 TSR; HOE 87.1678) [cited in BUA, 1997].
Steinberg S, Kaplan I (1984) The determination of low molecular weight aldehydes in rain, fog and mist by reversed phase liquid chromatography of the 2,4-dinitrophenylhydrazone derivatives. International Journal of Environmental and Analytical Chemistry, 18:253-266.
Suwa Y, Nagao M, Kosugi A, Sugimura T (1982) Sulfite suppresses the mutagenic property of coffee. Mutation Research, 102:383-391.
Takahashi M, Okamiya H, Furukawa F, Toyoda K, Sato H, Imaida K, Hayashi Y (1989) Effects of glyoxal and methylglyoxal administration on gastric carcinogenesis in Wistar rats after initiation with N-methyl-N'-nitro-N-nitrosoguanidine. Carcinogenesis, 10(10):1925-1927.
Thomas RG (1982) Volatilization from water. In: Lyman W, Reehl W, Rosenblatt D, eds. Handbook of chemical property estimation methods. Environmental behavior of organic compounds. New York, NY, McGraw-Hill, pp. 1-34.
Thornalley PJ (1993) The glyoxalase system in health and disease. Molecular Aspects of Medicine, 14(4):287-371.
Thornalley PJ (1995) Advances in glyoxalase research. Glyoxalase expression in malignancy, anti-proliferative effects of methylglyoxal, glyoxalase I inhibitor diesters and S-D-lactoylglutathione, and methylglyoxal-modified protein binding and endocytosis by advanced glycation endproduct receptor. Critical Reviews in Oncology/Hematology, 20(1-2):99-128.
Thornalley PJ (1998) Glutathione-dependent detoxification of alpha-oxoaldehydes by the glyoxalase system: involvement in disease mechanisms and antiproliferative activity of glyoxalase I inhibitors. Chemico-Biological Interactions, 111-112:137-151.
Thornalley PJ (2002) Glycation in diabetic neuropathy: characteristics, consequences, causes, and therapeutic options. International Review of Neurobiology, 50:37-57.
Thornalley PJ, McLellan AC, Lo TWC, Benn J, Sönksen PH (1996) Negative association of red blood cell reduced glutathione concentration with diabetic complications. Clinical Science, 91:575-582.
Thornalley PJ, Langborg A, Minhas HS (1999) Formation of glyoxal, methylglyoxal and 3-deoxyglucosone in the glycation of proteins by glucose. Biochemical Journal, 344:109-116.
Thornalley PJ, Yurek-George A, Argirov OK (2000) Kinetics and mechanism of the reaction of aminoguanidine with the alpha-oxoaldehydes glyoxal, methylglyoxal, and 3-deoxyglucosone under physiological conditions. Biochemical Pharmacology, 60(1):55-65.
Tichy FE, Bjorge T, Magnussen BF, Bengtsson PE, Mauss F (1998) Two-dimensional imaging of glyoxal (C2H2O2) in acetylene flames using laser-induced fluorescence. Applied Physics B, 66(1):115-119.
Tucker JD, Taylor RT, Christensen ML, Strout CL, Hanna ML, Carrano AV (1989) Cytogenetic response to 1,2-dicarbonyls and hydrogen peroxide in Chinese hamster ovary AUXB1 cells and human peripheral lymphocytes. Mutation Research, 224:269-279.
Ueno H, Segawa T, Hasegawa T, Nakamuro K, Maeda H, Hiramatsu Y, Okada S, Sayato Y (1991a) Subchronic oral toxicity of glyoxal via drinking water in rats. Fundamental and Applied Toxicology, 16:763-772.
Ueno H, Nakamuro K, Sayato Y, Okada S (1991b) Characteristics of mutagenesis by glyoxal in Salmonella typhimurium: contribution of singlet oxygen. Mutation Research, 251:99-107.
Ueno H, Nakamuro K, Sayato Y, Okada S (1991c) DNA lesion in rat hepatocytes induced by in vitro and in vivo exposure to glyoxal. Mutation Research, 260:115-119.
Ulrich P, Cerami A (2001) Protein glycation, diabetes, and aging. Recent Progress in Hormone Research, 56:1-21.
US EPA (1997) Exposure factors handbook. National Center for Environmental Assessment, US Environmental Protection Agency, August. Available at http://cfpub.epa.gov/ncea/cfm/recordisplay.cfm?deid=12464.
US EPA (1999) Determination of carbonyl compounds in drinking water by fast gas chromatography. Washington, DC, US Environmental Protection Agency, Office of Research and Development, September, pp. 1-38 (Method 556.1).
US EPA (2000) DermWin. Office of Pollution Prevention & Toxics, US Environmental Protection Agency. Available at http://www.epa.gov/oppt/.
Uter W, Schwanitz HJ, Lessmann H, Schnuch A (2001) Glyoxal is an important allergen for (medical care) cleaning staff. International Journal of Hygiene and Environmental Health, 204(4):251-253.
von der Hude W, Behm C, Gürtler R, Basler A (1988) Evaluation of the SOS chromotest. Mutation Research, 203:81-94.
Wangenheim J, Bolcsfoldi G (1988) Mouse lymphoma L5178Y thymidine kinase locus assay of 50 compounds. Mutagenesis, 3(3):193-205.
Wells-Knecht KJ, Zyzak DV, Litchfield JE, Thorpe SR, Baynes JW (1995) Mechanism of autoxidative glycosylation: identification of glyoxal and arabinose as intermediates in the autoxidative modification of proteins by glucose. Biochemistry (Washington), 34:3702-3709.
Whipple E (1970) The structure of glyoxal in water. Journal of the American Chemical Society, 92(24):7183-7186.
Whittaker M, Kersten PJ, Cullens D, Whittaker JW (1999) Identification of catalytic residues in glyoxal oxidase by targeted mutagenesis. Journal of Biological Chemistry, 274(51):36226-36232.
Witowski J, Korybalska K, Wisniewska J, Breborowicz A, Gahl GM, Frei U, Paalick-Deetjen J, Jörres A (2000) Effect of glucose degradation products on human peritoneal mesothelial cell function. Journal of the American Society of Nephrology, 11(4):729-739.
Wondrak GT, Cervantes-Laurean D, Roberts MJ, Qasem JG, Kim M, Jacobson EL, Jacobson MK (2002a) Identification of alpha-dicarbonyl scavengers for cellular protection against carbonyl stress. Biochemical Pharmacology, 63:361-373.
Xu GG, Yang CQ, Deng Y (2002) Applications of bifunctional aldehydes to improve paper wet strength. Journal of Applied Polymer Science, 83(12):2539-2547.
Yadav GD, Gupta VR (2000) Synthesis of glyoxalic acid from glyoxal. Process Biochemistry, 36:73-78.
Yamaguchi M, Ishida J, Xuan-Xuan Z, Nakamura M, Yoshitake T (1994) Determination of glyoxal, methylglyoxal, diacethyl, and 2,3-pentanedione in fermented foods by high-performance liquid chromatography with fluorescence detection. Journal of Liquid Chromatography, 17:203-211.
Yamaguchi T, Nakagawa K (1983) Mutagenicity of and formation of oxygen radicals by trioses and glyoxal derivatives. Agricultural and Biological Chemistry, 47(11):2461-2465.
Younes M (1997) Freie Radikale und Sauerstoffspezies. In: Marquardt H, Schäfer SG, eds. Lehrbuch der Toxikologie. Heidelberg, Spektrum Akademischer Verlag.
Zhou X, Mopper K (1990) Measurement of sub-parts-per-billion levels of carbonyl compounds in marine air by a simple cartridge trapping procedure followed by liquid chromatography. Environmental Science and Technology, 24:1482-1485.
Zimmermann F, Mohr A (1992) Formaldehyde, glyoxal, urethane, methyl carbamate, 2,3-butanedione, 2,3-hexanedione, ethyl acrylate, dibromoacetonitrile and 2-hydroxypropionitrile induce chromosome loss in Saccharomyces cerevisiae. Mutation Research, 270:151-166.
BUA (1997) [Glyoxal.] German Chemical Society (GDCh) Advisory Committee on Existing Chemicals of Environmental Relevance (BUA). Stuttgart, S. Hirzel, Wissenschaftliche Verlagsgesellschaft (BUA Report 187) (in German)
The objective of BUA assessments is to serve as a basis for the instigation of administrative measures when there are indications of risks of a chemical to health or to the environment.
For the BUA review process, the company that is in charge of writing the report (usually the largest manufacturer in Germany) prepares a draft report using literature from an extensive literature search as well as internal company studies. This draft is subject to a peer review in several readings of a working group consisting of representatives from government agencies, the scientific community, and industry.
The toxicological sections of this BUA report were prepared by Berufsgenossenschaft der Chemischen Industrie (BG Chemie, Toxicological Evaluations No. 177, 1996). The English version of the BUA report was published in 1998.
The draft CICAD on glyoxal was sent for review to IPCS national Contact Points and Participating Institutions, as well as to identified experts. Comments were received from:
R. Benson, Drinking Water Program, US Environmental Protection Agency, Denver, CO, USA
H.S. Chan, National Institute for Occupational Safety and Health, Cincinnati, OH, USA
R.S. Chhabra, National Institute of Environmental Health Sciences, Research Triangle Park, NC, USA
C. Cooke, Health and Safety Executive, Bootle, Merseyside, United Kingdom
P. Copestake, Toxicology Advice & Consulting Ltd, Sutton, United Kingdom
I. Desi, Department of Public Health, University of Szeged, Szeged, Hungary
J. Donohue, Office of Water, US Environmental Protection Agency, Washington, DC, USA
C. Elliot-Minty, Health and Safety Executive, Bootle, Merseyside, United Kingdom
L. Fishbein, Private consultant, Fairfax, VA, USA
E. Frantik, National Institute of Public Health, Prague, Czech Republic
R. Gatehouse, Environment Australia, Canberra, Australia
T. Gebhart, BASF Aktiengesellschaft, Jockgrim, Germany
P. Harvey, Department of Health and Ageing, Sydney, Australia
R.F. Hertel, Federal Institute for Risk Assessment, Berlin, Germany
P. Joseph, National Institute for Occupational Safety and Health, Morgantown, WV, USA
R. Morgenstern, Karolinska Institute, Stockholm, Sweden
T.-M. Ong, National Institute for Occupational Safety and Health, Morgantown, WV, USA
V. Riihimäki, Finnish Institute of Occupational Health, Helsinki, Finland
J.L. Stauber, CSIRO Energy Technology, Bangor, NSW, Australia
K. Ziegler-Skylakakis, European Commission, Luxembourg
Varna, Bulgaria
8-11 September 2003
Members
Dr I. Benchev, Sofia, Bulgaria
Dr R. Chhabra, National Institute of Environmental Health Sciences, Research Triangle Park, NC, USA
Dr C. De Rosa, Agency for Toxic Substances and Disease Registry, Centers for Disease Control and Prevention, Atlanta, GA, USA
Dr S. Dobson, Centre for Ecology and Hydrology, Monks Wood, Abbots Ripton, Huntingdon, Cambridgeshire, United Kingdom
Dr G. Dura, National Institute of Environment, József Fodor Public Health Centre, Budapest, Hungary
Dr L. Fishbein, Fairfax, VA, USA
Dr H. Gibb, National Center for Environmental Assessment, US Environmental Protection Agency, Washington, DC, USA
Dr R.F. Hertel, Federal Institute for Risk Assessment, Berlin, Germany
Mr P. Howe, Centre for Ecology and Hydrology, Monks Wood, Abbots Ripton, Huntingdon, Cambridgeshire, United Kingdom
Dr S. Ishimitsu, Division of Safety Information on Drug, Food and Chemicals, National Institute of Hygienic Sciences, Tokyo, Japan
Dr D. Kanungo, Central Insecticides Board, Directorate of Plant Protection, Quarantine & Storage, Ministry of Agriculture, Haryana, India
Dr J. Kielhorn, Fraunhofer Institute for Toxicology and Experimental Medicine, Hanover, Germany
Ms B. Meek, Environmental Health Directorate, Health Canada, Ottawa, Ontario, Canada
Dr T. Morita, Division of Safety Information on Drug, Food and Chemicals, National Institute of Hygienic Sciences, Tokyo, Japan
Mr F.K. Muchiri, Directorate of Occupational Health and Safety Services, Nairobi, Kenya
Dr L. Olsen, Biological Monitoring & Health Assessment Branch, Division of Applied Research & Technology, National Institute for Occupational Safety and Health, Cincinnati, OH, USA
Dr N. Rizov, National Center of Hygiene, Medical Ecology and Nutrition, Sofia, Bulgaria
Dr P. Schulte, Education and Information Division, National Institute for Occupational Safety and Health, Cincinnati, OH, USA
Dr J. Sekizawa, Faculty of Integrated Arts and Sciences, Tokushima University, Tokushima, Japan
Dr F. Petrova Simeonova, Sofia, Bulgaria
Dr S. Soliman, Faculty of Agriculture, Alexandria University, El Shatby, Alexandria, Egypt
Dr J. Stauber, CSIRO Energy Technology, Centre for Advanced Analytical Chemistry, Bangor, NSW, Australia
Mr P. Watts, Toxicology Advice & Consulting Ltd, Surrey, United Kingdom
Ms D. Willcocks, National Industrial Chemicals Notification and Assessment Scheme, Sydney, NSW, Australia
Dr K. Ziegler-Skylakakis, European Commission, Luxembourg
Observers
Dr S. Jacobi, Degussa AG, Fine Chemicals, Hanau-Wolfgang, Germany
Mr M. Southern, Shell International Petroleum Company Ltd, London, United Kingdom
Dr W. ten Berge, DSM, Heerlen, The Netherlands
Secretariat
Dr A. Aitio, International Programme on Chemical Safety, World Health Organization, Geneva, Switzerland
Mr T. Ehara, International Programme on Chemical Safety, World Health Organization, Geneva, Switzerland
|
AGE |
advanced glycation end-product |
|
BOD |
biochemical oxygen demand |
|
CAS |
Chemical Abstracts Service |
|
CHO |
Chinese hamster ovary |
|
CICAD |
Concise International Chemical Assessment Document |
|
CML |
Nε-(carboxymethyl)lysine |
|
dC |
deoxycytidine |
|
dG |
deoxyguanosine |
|
DNA |
deoxyribonucleic acid |
|
DNPH |
dinitrophenylhydrazine |
|
EC50 |
median effective concentration |
|
ECD |
electron capture detection |
|
EHC |
Environmental Health Criteria |
|
FPG |
formamidopyrimidine N-glycosylase |
|
GC |
gas chromatography |
|
GSH |
glutathione |
|
GST-P |
glutathione-S-transferase placental form |
|
HPLC |
high-performance liquid chromatography |
|
ICSC |
International Chemical Safety Card |
|
ILO |
International Labour Organization |
|
IPCS |
International Programme on Chemical Safety |
|
Koc |
soil sorption coefficient |
|
Kow |
octanol/water partition coefficient |
|
Kp |
permeability coefficient from water |
|
LC50 |
median lethal concentration |
|
LD50 |
median lethal dose |
|
LOAEL |
lowest-observed-adverse-effect level |
|
LOEL |
lowest-observed-effect level |
|
MS |
mass spectrometry |
|
MW |
molecular weight |
|
NOAEL |
no-observed-adverse-effect level |
|
NOEC |
no-observed-effect concentration |
|
NOEL |
no-observed-effect level |
|
OECD |
Organisation for Economic Co-operation and Development |
|
PEC |
predicted environmental concentration |
|
PFBHA |
pentafluorobenzyl hydroxylamine |
|
PIM |
Poison Information Monograph |
|
PNEC |
predicted no-effect concentration |
|
SI |
International System of Units (Système international d'unités) |
|
ThOD |
theoretical oxygen demand |
|
UNEP |
United Nations Environment Programme |
|
USA |
United States of America |
|
UV |
ultraviolet |
|
WHO |
World Health Organization |
In section 6.2.2, the exposure to a glyoxal-containing aerosol disinfectant was estimated for the scenario of a stable. The deterministic model for predicting aerosol exposure and inhalation during the spray application of biocidal products, which was developed by Prof. W. Koch, Fraunhofer Institute for Toxicology and Experimental Medicine, Hanover, was employed for the calculation. This model (Droplet Simulation Model) is mentioned in the 2002 version of Technical Notes for Guidance. Human Exposure to Biocidal Products. Guidance on Exposure Estimation. Final June 2002 (CA-Jul02-Doc.7.2-Part 2, Chapter 3.4, p. 225; for European Commission, DG Environment).
Assumptions for the model calculation performed
|
Sprayer |
Type of sprayer |
Type of nozzle |
Spraying conditions |
Aerosol diametera |
|
Frowein "Spray Boss" |
air cushion sprayer |
large, fan-shaped nozzle |
2000 hPa; |
304 µm (d50) |
a For droplet distribution, dx means that x percentile have given aerosol diameter.
Exposure concentrations and doses
Mean exposure concentration (mg/m³)
|
Aerosol characteristics |
Exposure time |
|
|
6 min |
15 min |
|
|
Able to pass into the alveolar space (<4.5 µm) |
0.012 |
0.016 |
|
Able to pass into the thorax (<10 µm) |
0.023 |
0.031 |
|
Total inhalable |
0.024 |
0.032 |
Mean inhaled dose (mg)
|
Aerosol characteristics |
Exposure time |
|
|
6 min |
15 min |
|
|
Able to pass into the alveolar space (<4.5 µm) |
0.0008 |
0.0025 |
|
Able to pass into the thorax (<10 µm) |
0.0015 |
0.0045 |
|
Total inhalable |
0.0016 |
0.0048 |
Le présent CICAD consacré au glyoxal a été préparé par l'Institut Fraunhofer de toxicologie et médecine expérimentale de Hanovre (Allemagne). Il s'inspire de rapports établis par le Comité consultatif allemand sur les substances chimiques d'importance écologique (BUA, 1997). Il a été procédé à un dépouillement bibliographique exhaustif des bases de données correspondantes jusqu'en février 2003, afin de rechercher toute référence intéressante à des publications postérieures à celles qui sont prises en compte dans les rapports en question. Des informations sur la préparation et l'examen par des pairs des sources documentaires utilisées sont données à l'appendice 1. L'appendice 2 fournit des renseignements sur l'examen par des pairs du présent CICAD. Ce CICAD a été examiné et approuvé en tant qu'évaluation internationale lors d'une réunion du Comité d'évaluation finale qui s'est tenue à Varna (Bulgarie) du 8 au 11 septembre 2003. La liste des participants à cette réunion figure à l'appendice 3. La fiche internationale sur la sécurité chimique du glyoxal (ICSC 1162), établie par le Programme international sur la sécurité chimique (IPCS, 2002), est également reproduite dans le présent document.
Le glyoxal anhydre (No CAS
Les principaux compartiments environnementaux dans lesquels se retrouve le glyoxal sont l'hydrosphère et le sol (respectivement dans la proportion de 46 et 54 %). Il est également présent dans l'air, mais dans une moindre mesure (< 1 %). La concentration de glyoxal relevée dans l'air ambiant aux Etats-Unis, en Europe et en Asie va d'environ 0,1 à 10 μg/m3. Dans les cours d'eau et les eaux souterraines d'Europe, on fait état de concentrations pouvant aller jusqu'à 12 μg/litre. Le glyoxal est un sous-produit de la désinfection par l'ozone et on en a mis en évidence dans de l'eau de boisson à des concentrations de quelques μg par litre.
On constate souvent, dans les denrées alimentaires et les boissons fermentées, la présence de glyoxal résultant de l'activité microbienne, de l'autooxydation non enzymatique de l'huile ou encore des réactions de brunissement des sucres. Dans différentes marques de bière et dans des vins de divers crus, de même que dans d'autres boissons comme le thé, on en a décelé la présence à des concentrations allant de d'environ 20 μg/litre (thι noir) ΰ 1556 μg/litre (vin de Xιrès). Par ailleurs, on en a trouvé dans plusieurs produits alimentaires comme la pâte de soja et le yoghourt (0,63 à 4,2 mg/kg), dans des produits de boulangerie comme le pain (0,07 à 1,6 mg/kg) et divers produits végétaux (3-14 mg/kg) ou huiles comestibles (jusqu'à 6,5 mg/kg).
Le glyoxal libéré dans l'environnement subit une transformation rapide sous l'effet de processus abiotiques, dans lesquels interviennent notamment des radicaux hydroxyles produits par voie photochimique. En raison de son faible coefficient de sorption aux particules du sol (Koc), le composé est susceptible de passer par lessivage du sol aux eaux souterraines. Il est toutefois rapidement biodégradé et les enzymes bactériennes ou fongiques le transforment en peu de temps. Comme son coefficient de partage entre l'octanol et l'eau (Kow) est peu élevé, il n'a vraisemblablement guère tendance à subir une bioaccumulation.
Les principales voies d'exposition professionnelle au glyoxal lors de son utilisation comme désinfectant consistent dans l'inhalation d'aérosols ou dans l'absorption transcutanée du produit. L'exposition de la population dans son ensemble est principalement due à la consommation de denrées alimentaires contenant du glyoxal, mais elle peut également résulter d'une forte pollution atmosphérique ou de la présence de traces de glyoxal dans l'eau de boisson.
Le métabolisme cellulaire normal entraîne la production de glyoxal endogène par toutes sortes de voies enzymo-indépendantes. Le glyoxal endogène résulte également de la métabolisation et de l'oxydation microsomique d'autres molécules comme le glycolaldéhyde, l'éthylène-glycol, et les beta-hydroxy-N-nitrosamines. La concentration du glyoxal dans le plasma humain serait de 0,1 à 1 μmol/litre, des concentrations plus élevées étant observées chez les diabétiques et les insuffisants rénaux. Dans les produits biologiques, moins de 10 % du glyoxal présent se trouve en solution aqueuse sous forme non liée (glyoxal libre et hydrates), car la majorité des groupements carbonyles réactifs sont réversiblement liés aux résidus cystéinyle, lysyle et argininyle des protéines.
Le glyoxal, qui attaque les groupements amino des protéines, des nucléotides et des lipides, est considéré comme un important intermédiaire dans la formation des produits de la glycation avancée (AGE). La glycation avancée altère la fonction des protéines et inactive les enzymes, avec pour conséquence des troubles du métabolisme cellulaire, une mauvaise protéolyse ainsi qu'une inhibition de la prolifération cellulaire et de la synthèse protéique. Les effets délétères du glyoxal sont combattus par un système enzymatique ubiquiste, celui de la glyoxalase glutathion (GSH)-dépendante qui transforme le glyoxal en glycolate moins réactif.
La toxicité aiguë du glyoxal pour les animaux de laboratoire est faible à modérée, selon la concentration effective du composé dans le produit étudié. Chez le rat soumis pendant 4 h à l'inhalation d'un aérosol à 40 % de glyoxal, la CL50 est de 2440 mg/m3; la DL50 par voie orale varie de 3000 à 9000 mg/kg de poids corporel (avec une sensibilité plus marquée chez les femelles), la DL50 cutanée étant supérieure à 2000 mg/kg de poids corporel. Après exposition par la voie respiratoire, l'effet prédominant est une irritation des yeux et des voies aériennes, accompagnée d'une hyperémie et d'une sécrétion spumeuse au niveau pulmonaire. Une exposition par voie orale détermine des effets macroscopiques tels qu'une irritation et une congestion des voies digestives, la congestion s'étendant également au poumon, au rein et aux surrénales. Dans les principaux organes cibles, à savoir le pancréas et le rein, l'action toxique du glyoxal conduit à de graves lésions dégénératives évoquant celles que provoque le diabète.
Des études au cours desquelles on a fait inhaler du glyoxal à des rats pendant une courte période (29 jours), montrent que la dose sans effet observable au niveau du larynx (NOEL) est de 0,6 mg/m3 (concentration nominale égale à 0,4 mg/m3), la dose sans effets généraux observables (contrôle du poids corporel, des paramètres hématologiques et biochimiques, analyse d'urine, examen macroscopique et histologique) étant supérieure à 8.9 mg/m3 (concentration nomimale égale à 10 mg/m3). Une étude de 28 jours pendant laquelle du glyoxal a été administré à des rats dans leur eau de boisson a permis de fixer à 100 mg/kg p.c. par jour la dose de composé sans effet nocif observable (NOAEL). Pour d'autres rats qui en avaient reçu pendant 90 jours dans leur alimentation, la NOAEL s'est révélée égale à 125 mg/kg de poids corporel par jour (dose correspondant à 100 % de glyoxal). Les effets observés dans ces deux dernières études aux doses les plus élevées consistaient en une diminution de la consommation d'eau et de nourriture (uniquement dans la première étude) et en un ralentissement du gain de poids (dans les deux études). Lors d'une autre étude portant sur des points d'aboutissement plus sensibles de l'action toxique (paramètres biochimiques sériques), la plus faible dose étudiée chez le rat — à savoir 107 mg/kg p.c. par jour (soit 99 % de glyoxal) — correspondait à la dose la plus faible provoquant un effet nocif observable (LOAEL) obtenue lors d'une exposition de 90 jours par l'intermédiaire de l'eau de boisson. Une étude de 90 jours sur des chiens exposés par la voie alimentaire n'a révélé aucune anomalie attribuable au composé à la dose journalière maximale de 115 mg/kg p.c. (dose correspondant à 100 % de glyoxal).
L'expérimentation animale montre que des solutions à 30 et 40 % de glyoxal provoquent une irritation cutanée légère à marquée, selon la durée de contact. Le glyoxal est irritant pour les muqueuses et il se comporte comme un sensibilisateur cutané chez l'Homme et les animaux de laboratoire.
Des effets foetotoxiques n'ont été observés qu'à des doses qui étaient elles-mêmes toxiques pour la mère. Les études toxicologiques portant sur le développement du rat ont permis de fixer à > 300 mg de dihydrate de glyoxal par kg de poids corporel la dose journalière sans effet embryotoxique observable (NOEL) — soit > 185 mg de glyoxal par kg de poids corporel et par jour) — alors que la dose la plus faible provoquant un effet observable chez la mère (LOEL) (réduction du gain de poids) était de 200 mg de ce même dihydrate par kg p.c.par jour (soit 123 mg de glyoxal par kg p.c. par jour). Des études sur le lapin destinées à déterminer l'intervalle de toxicité pour le développement ont permis d'obtenir une NOEL de 200 mg de dihydrate de glyoxal par kg p.c. par jour
(soit 123 mg/kg p.c. par jour) tant pour la toxicité maternelle que pour la toxicité embryonnaire.
In vitro, le glyoxal présente une génotoxicité directe pour les cellules bactériennes et mammaliennes et provoque notamment la formation d'adduits avec l'ADN, des mutations, des aberrations chromosomiques, une réparation de l'ADN, des échanges entre chromatides soeurs et des ruptures monocaténaires de l'ADN. In vivo, on a mis en évidence l'activité génotoxique du glyoxal au niveau de son point d'application sur la muqueuse pylorique du rat, par l'observation d'une synthèse non programmée et de ruptures monocaténaires de l'ADN. Après administration par voie orale à des rats, on a également observé la rupture des brins de l'ADN dans les hépatocytes. Il n'y a pas eu d'étude de cancérogénicité in vivo chez des animaux exposés au glyoxal par la voie respiratoire. Sur un modèle de cancérogénèse en deux étapes (paroi glandulaire de l'estomac du rat Wistar), on a constaté que le glyoxal se comportait comme un promoteur tumoral. Par contre, il s'est montré inactif lors d'un test à court terme consistant dans la recherche de foyers hépatiques positifs. Une étude destinée à mettre en évidence l'activité tumoro-promotrice du glyoxal par des tests de transformation cutanée et cellulaire, a donné des résultats négatifs.
En s'appuyant sur l'étude d'inhalation de 29 jours qui donne une NOEL de 0,6 mg/m3 chez le rat pour les effets localisés au larynx et en appliquant un premier coefficient d'incertitude de 10 pour tenir compte des différences interespèces et un second de 10 également pour les différences interindividuelles, on arrive à une concentration tolérable estimative de 6 μg/m3 pour les effets localisés au larynx en cas d'exposition de brève durée.
Pour établir une évaluation représentative du risque couru par la population dans son ensemble, on a imaginé un scénario d'exposition correspondant au cas le plus grave. En supposant que la dose journalière ingérée par voie alimentaire correspond à un maximum de 10 mg de glyoxal, on obtient une dose estimative de 0,16 mg de glyoxal par jour et par kg de poids corporel. Cette valeur est du même ordre que la dose journalière tolérable d'environ 0,2 mg par kg de poids corporel déterminée pour une exposition par voie orale pendant toute la durée de l'existence.
Dans une seconde évaluation du risque couru cette fois par une infirmière, un membre du personnel de nettoyage d'un hôpital ou un particulier utilisant un désinfectant à base de glyoxal, on a supposé que ces personnes se servaient d'un produit ordinaire contenant 7,5 g de composé pour 100 g — soit 7,5 % de glyoxal — ramené à 0,075 % de glyoxal par dilution 1/100ème pour désinfecter ou nettoyer certaines surfaces. En arrondissant à 0,1 % la teneur de cette solution et en utilisant un modèle de calcul, on arrive à une dose journalière absorbée d'environ 4 μg/kg p.c., le poids corporel moyen étant pris égal à 64 kg. C'est donc une valeur beaucoup plus faible (environ 50 fois) que la dose tolérable journalière d'environ 0,2 mg/kg p.c. relative à une exposition sur toute la durée de la vie. Cependant, en prenant le cas le plus grave, c'est-à-dire une exposition à une solution à 4 % de glyoxal et en partant des mêmes hypothèses que précédemment, on obtiendrait une dose absorbée d'environ 0,15 mg/kg p.c., ce qui correspond approximativement à la dose journalière tolérable d'environ 0,2 mg/kg p.c. pour une exposition pendant toute la durée de la vie.
Pour la dernière évaluation du risque, on a pris à titre d'exemple le cas d'un cultivateur qui pulvérise un produit biocide à base de glyoxal pour désinfecter une étable. En tenant compte des hypothèses formulées, le modèle de calcul utilisé donne, pour cette personne, une exposition de brève durée à 24 μg de glyoxal par m3 pendant 6 minutes et à 32 μg de glyoxal par m3 pendant 15 minutes. Ces valeurs sont à comparer à la concentration tolérable estimative de 6 μg/m3 pour l'irritation du larynx provoquée par une exposition de brève durée. Dans le cas présenté ici de l'utilisation de glyoxal en pulvérisations désinfectantes, il y a perception d'un risque d'effets irritants au niveau du larynx.
On a montré que le glyoxal inhibe l'activité des bactéries aérobies et anaérobies, des algues vertes (CE50 à 96 h égale à environ 149 mg/litre pour Pseudokirchneriella subcapitata — anciennement Selenastrum capricornutum — et d'une espèce d'invertébré. Sur les quatre espèces de poissons étudiées, c'est Pimephales promelas qui présentait la valeur de la CL50 à 96 h la plus faible, soit 215 mg/litre.
Afin de caractériser de manière représentative le risque pour l'environnement aquatique, on a calculé le rapport de la concentration environnementale prévisible localement (PEC), déterminée sur la base de mesures récentes, à la concentration correspondante sans effet prévisible (PNEC). La valeur de la PNEC a été estimée à 149 μg/litre pour les eaux superficielles en appliquant un coefficient d'incertitude de 1000 à la valeur la plus faible de la CE50, estimée à 149 mg/litre. Pour obtenir le rapport PEC/PNEC, on a pris la valeur la plus élevée de la concentration en glyoxal trouvée dans les eaux superficielles (1,9 μg/litre), ce qui donne un rapport de 0,013. Comme cette valeur est infιrieure ΰ 1, il n'y a pas lieu d'obtenir d'autres données, ni de pousser plus loin l'expérimentation ou les mesures de réduction du risque.
La seule étude disponible a permis d'obtenir une valeur de 68 mg/litre pour la concentration sans effet observable (NOEC) sur Helianthus tuberosus, l'effet retenu étant l'inhibition de la prolifération de fragments de rhizome. La valeur correspondante de la CE50 était de 136 mg/litre. Comme on ne dispose d'aucune autre donnée toxicologique caractéristique concernant les effets toxiques du glyoxal sur les microorganismes ou les invertébrés terrestres, on n'a pas été en mesure de caractériser de manière fiable et quantitative le risque auquel ces organismes sont exposés.
El presente CICAD sobre el glioxal fue preparado por el Instituto Fraunhofer de Toxicología y Medicina Experimental de Hannover (Alemania). Se basa en los informes compilados por el Comité Consultivo Alemán sobre las Sustancias Químicas Importantes para el Medio Ambiente (BUA, 1997). Se realizó una búsqueda bibliográfica amplia en las bases de datos pertinentes hasta febrero de 2003 para encontrar cualquier referencia publicada después de las incorporadas a esos informes. La información relativa a la preparación y el examen colegiado del documento original se presenta en el apéndice 1. La información sobre el examen colegiado de este CICAD aparece en el apéndice 2. Este CICAD se aprobó como evaluación internacional en una reunión de la Junta de Evaluación Final, celebrada en Varna (Bulgaria) del 8 al 11 de septiembre de 2003. La lista de participantes en esta reunión figura en el apéndice 3. También se reproduce en este documento la Ficha internacional de seguridad química (ICSC 1162) para el glioxal, preparada por el Programa Internacional de Seguridad de las Sustancias Químicas (IPCS, 2002).
El glioxal anhidro (CAS Nº
Los compartimentos destinatarios predominantes del glioxal en el medio ambiente son la hidrosfera y el suelo (con alrededor del 46% y el 54%, respectivamente) y en menor medida el aire (< 1%). Las concentraciones notificadas de glioxal en el aire atmosférico de los Estados Unidos, Europa y Asia oscilan entre alrededor de 0,1 y 10 µg/m3. En los ríos y las aguas freáticas europeos se han notificado concentraciones de hasta 12 µg/l. El glioxal es un subproducto de la desinfección con ozono y en el agua de bebida se han detectado concentraciones tan sólo del orden de µg/l.
Debido a la actividad microbiana, así como a la autooxidación no enzimática del aceite o las reacciones de caramelización de los sacáridos, es frecuente detectar glioxal en los alimentos y bebidas fermentados. Se encontró en diferentes marcas de cerveza, vino y otros bebidas, como el té, en concentraciones que variaban entre unos 20 µg/l (té negro) y 1556 µg/l (vino de jerez). Además, se detectó en una serie de productos fermentados, como la pasta de soja y el yogur (0,63-4,2 mg/kg), productos de panadería como el pan (0,07-1,6 mg/kg), distintos materiales vegetales (3-14 mg/kg) y aceites comestibles (hasta 6,5 mg/kg).
El glioxal liberado en el medio ambiente se transforma con rapidez mediante procesos abióticos, como la acción de radicales hidroxilo producidos por vía fotoquímica. Debido al bajo coeficiente de sorción en el suelo (Koc) notificado para este compuesto, es posible su lixiviación hacia el agua freática. Sin embargo, se biodegrada con facilidad y sufre una transformación enzimática rápida por la acción de bacterias y hongos. El bajo valor de su log del coeficiente de reparto octanol/ agua (Kow) indica que es poco probable la bioacumulación de glioxal.
Las principales vías de exposición profesional al glioxal durante su utilización como desinfectante son por inhalación de aerosoles o absorción cutánea. La exposición de la población general se debe principalmente al consumo de alimentos que contienen glioxal, pero también se podría producir mediante el aire contaminado de regiones urbanas y la presencia de cantidades ínfimas en el agua de bebida.
El glioxal se produce de manera endógena durante el metabolismo celular normal mediante numerosas vías enzimáticas independientes. Es también un producto del metabolismo y la oxidación microsómica de otros compuestos, como el glicolaldehído, el etilenglicol y las N-nitrosaminas beta-hidroxi sustituidas. Se han notificado concentraciones de glioxal en el plasma sanguíneo humano de 0,1-1 µmoles/l, con niveles más elevados en pacientes con diabetes o insuficiencia renal. En los materiales biológicos, la concentración de glioxal presente en forma no combinada en solución acuosa es inferior al 10% (glioxal libre e hidratos), puesto que la mayor parte de los grupos carbonilo reactivos están unidos con carácter reversible a residuos de cisteína, lisina y arginina de las proteínas.
Se considera que el glioxal, que ataca los grupos amino de las proteínas, los nucleótidos y los lípidos, es un intermediario importante en la formación de productos finales de la glicación avanzada. La modificación de estos productos altera la función de las proteínas e inactiva las enzimas, produciendo trastornos del metabolismo celular, alteraciones de la proteolisis e inhibición de la proliferación celular y la síntesis de proteínas. Los efectos perjudiciales del glioxal en estado muy reactivo se contrarrestan mediante un sistema ubicuo de glioxalasa dependiente del glutatión, que transforma el glioxal en glicolato, menos reactivo.
La toxicidad aguda del glioxal en animales de experimentación es de baja a moderada, en función de la concentración real de glioxal en el producto que se examina. En ratas, la CL50 del glioxal al 40% para una inhalación única de aerosol de cuatro horas es de 2440 mg/m3, el valor de la DL50 por vía oral oscila entre 3000 y 9000 mg/kg de peso corporal (con sensibilidad más elevada en las hembras) y los valores de la DL50 por vía cutánea son >2000 mg/kg de peso corporal. Tras la exposición por inhalación predominan las irritaciones locales de los ojos y los órganos respiratorios, así como hiperemia y secreción espumosa en los pulmones. Las observaciones macroscópicas después de la exposición oral al glioxal son de irritación del tracto gastrointestinal y congestión del tracto gastrointestinal, los pulmones, el riñón y las glándulas suprarrenales. En los órganos destinatarios principales, el páncreas y el riñón, la acción tóxica del glioxal provoca cambios degenerativos graves semejantes a los inducidos durante la diabetes.
En los estudios de exposición breve (29 días) de ratas al glioxal por inhalación se puso de manifiesto una concentración sin efectos observados (NOEL) de 0,6 mg/m3 (la concentración nominal fue de 0,4 mg/m3) para los efectos locales en la laringe y una NOEL de >8,9 mg/m3 (la concentración nominal fue de 10 mg/m3) para los efectos sistémicos (examen del peso corporal, parámetros hematológicos y bioquímicos, análisis de orina, examen macroscópico e histológico). En un estudio de 28 días en el que se administró a ratas glioxal en el agua de bebida se obtuvo una concentración sin efectos adversos observados (NOAEL) de 100 mg de glioxal/kg de peso corporal al día. La administración de glioxal a ratas con los alimentos durante 90 días produjo una NOAEL de 125 mg/kg de peso corporal al día (siendo la dosis de glioxal del 100%). Los efectos con dosis más altas notificados en estos dos últimos estudios fueron una reducción de la ingesta de agua y de alimentos (sólo en el primer estudio) y retraso del aumento del peso corporal (en ambos estudios). En un estudio en el que se examinaban los efectos finales más sensibles (bioquímica clínica del suero), la dosis más baja sometida prueba de 107 mg/kg de peso corporal al día (glioxal al 99%) correspondió a la concentración más baja con efectos adversos observados (LOAEL) para una exposición de ratas durante 90 días en el agua de bebida. En un estudio de alimentación de 90 días con perros no se observó ningún cambio relacionado con la sustancia al utilizar la dosis máxima de 115 mg/kg de peso corporal al día (dosis correspondiente a glioxal al 100%).
En estudios con animales, el glioxal acuoso al 30% y el 40% provocó irritaciones cutáneas entre ligeras y manifiestas, en función del tiempo de aplicación. El glioxal es irritante de las membranas mucosas y actúa como agente sensibilizador cutáneo en las personas y los animales de experimentación.
Sólo se produjeron efectos fetotóxicos con dosis de glioxal que inducían toxicidad materna. En estudios de toxicidad en el desarrollo con ratas, la NOEL para la embriotoxicidad fue >300 mg de dihidrato de glioxal/kg de peso corporal al día (correspondientes a >185 mg de glioxal/kg de peso corporal al día), mientras que la concentración más baja con efectos observados (LOEL) (disminución del aumento de peso corporal) para la toxicidad materna fue de 200 mg de dihidrato de glioxal/kg de peso corporal al día (correspondientes a 123 mg de glioxal/kg de peso corporal al día). Los estudios para determinar las normas de toxicidad en el desarrollo en conejos dieron una NOEL de 200 mg de dihidrato de glioxal/kg de peso corporal al día (correspondientes a 123 mg de glioxal/kg de peso corporal al día) para la toxicidad materna y la embriotoxicidad.
El glioxal es directamente genotóxico in vitro en células de bacterias y de mamíferos, induciendo, por ejemplo, aductos de ADN, mutaciones, aberraciones cromosómicas, reparación de ADN, intercambio de cromátidas hermanas y roturas de cadenas sencillas de ADN. In vivo se estableció una actividad genotóxica del glioxal en el lugar de aplicación en la mucosa pilórica de ratas mediante la demostración de síntesis de ADN no programada y roturas de cadenas sencillas de ADN. Tras la aplicación oral se observó también rotura de cadenas de ADN en el hígado de la rata. No hay biovaloraciones de la carcinogénesis con la exposición por inhalación al glioxal. El glioxal mostró una actividad de inducción de tumores en un modelo de carcinogénesis gástrica glandular en dos etapas con ratas Wistar machos, mientras que fue inactivo en una valoración breve de focos del hígado. En una valoración de la actividad de inducción de tumores del glioxal en la piel y en valoraciones de transformación celular se obtuvieron resultados negativos.
Tomando el estudio de inhalación de 29 días en ratas expuestas al glioxal, en el que se obtuvo una NOEL de 0,6 mg/m3 para efectos locales en la laringe y utilizando factores de incertidumbre de 10 para diferencias interespecíficas y 10 para las interindividuales, se estimó que la concentración tolerable para los efectos locales en la laringe durante una exposición breve era de 6 µg/m3.
En una evaluación del riesgo de muestra para la población general se ha compilado un modelo de exposición como hipótesis del caso peor. Utilizando la ingesta diaria máxima de 10 mg de glioxal con los alimentos, se puede calcular una cantidad estimada de 0,16 mg de glioxal/kg de peso corporal al día. Esta es semejante a la ingesta tolerable de unos 0,2 mg/kg de peso corporal al día para la exposición oral al glioxal durante toda la vida.
En una segunda evaluación del riesgo de muestra, para una enfermera, una persona encargada de la limpieza en un hospital o un consumidor que utilice desinfectante, se utiliza una marca común de desinfectante (7,5 g en 100 g = 7,5% de glioxal) con una dilución del 1% para la desinfección y limpieza de superficies (es decir, 0,075% de glioxal). Utilizando el redondeo a una solución de glioxal del 0,1% y un cálculo derivado de un modelo se obtiene una ingesta de unos 4 µg/kg de peso corporal al día, suponiendo un peso corporal de 64 kg. Este valor es muy inferior (50 veces) a la ingesta tolerable de unos 0,2 mg/kg de peso corporal al día para la exposición por vía oral durante toda la vida. Sin embargo, utilizando una exposición en el caso peor a un 4% de glioxal y las mismas hipótesis expuestas más arriba se obtendría una ingesta de alrededor de 0,15 mg/kg de peso corporal, que es aproximadamente igual a la dosis diaria tolerable de unos 0,2 mg/kg de peso corporal al día para la exposición por vía oral durante toda la vida.
En la evaluación final del riesgo de muestra, se utilizó como ejemplo el caso de un agricultor que aplicaba productos biocidas con glioxal mediante pulverización para desinfectar un establo. El cálculo del modelo utilizando las hipótesis establecidas pronostica una exposición breve a una concentración de 24 µg de glioxal/m3 en seis minutos y de 32 µg de glioxal/m3 en 15. Esto se puede comparar con la concentración tolerable estimada de 6 mg/m3 para los efectos locales en la laringe tras una exposición breve. Se detecta un riesgo de efectos locales en la laringe y de irritación cutánea debido a la aplicación de glioxal mediante pulverización.
Se ha demostrado que la exposición al glioxal inhibe las actividades de bacterias tanto aerobias como anaerobias, de algas verdes (valor de la CE50 a las 96 horas de unos 149 mg/l para Pseudokirchneriella subcapitata [antes conocida como Selenastrum capricornutum]) y de una especie de invertebrado. En cuatro especies de peces sometidos a prueba, el valor más bajo notificado de la CL50 a las 96 horas fue de 215 mg/l (Pimephales promelas).
Se realizó una caracterización del riesgo de muestra para el medio acuático calculando la razón entre una concentración local prevista en el medio ambiente (PEC), basándose en datos medidos en fecha reciente, y la concentración prevista sin efectos (PNEC) correspondiente. Se estimó una PNEC de 149 µg/l para el agua superficial a partir del valor más bajo de la CE50 de 149 mg/l, aplicando un factor de incertidumbre de 1000. Con la concentración más alta de glioxal medida recientemente en el agua superficial (1,9 µg/l), se obtuvo un cociente PEC/PNEC de 0,013. Como este valor es inferior a 1, no hay necesidad de más información, pruebas o medidas de reducción del riesgo.
En el único estudio disponible se determinó una concentración sin efectos observados (NOEC) de 68 mg/l para la inhibición de la proliferación de fragmentos de rizoma de Helianthus tuberosus, con un valor correspondiente de la CE30 de 136 mg/l. Dado que no se dispone de datos adicionales para la caracterización de los efectos tóxicos del glioxal en los microorganismos o invertebrados terrestres, no es posible realizar una caracterización cuantitativa del riesgo fiable.
ENDNOTES:
See Also:
Toxicological Abbreviations